
 Lens - nKaryoTyping: Semantic Chromosome Inheritance Across Layers
Lens - nKaryoTyping: Semantic Chromosome Inheritance Across Layers
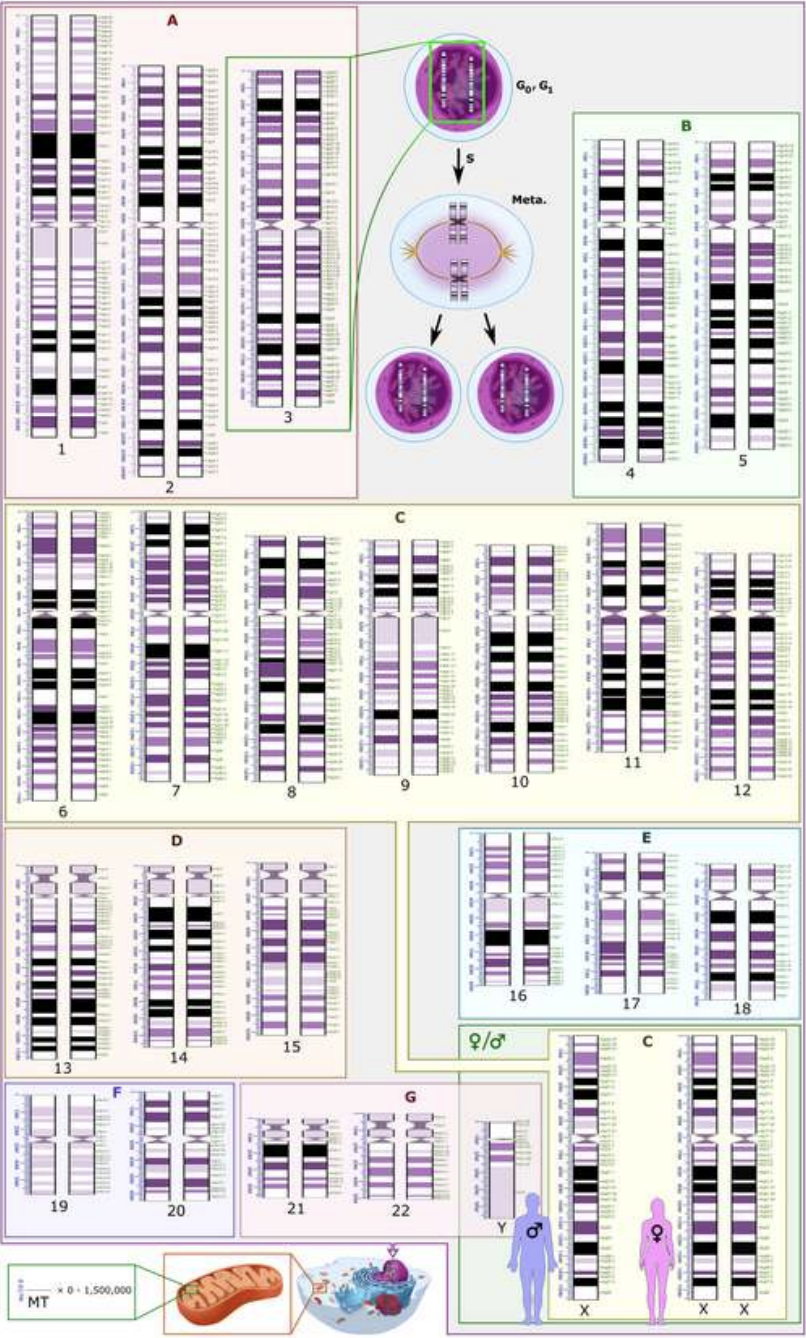
As foundation models grow in complexity and cultural diversity, the need to understand semantic inheritance across neural layers becomes increasingly paramount. To this end, we introduce nKaryotyping – a conceptual and visual framework inspired by biological karyotyping, which charts chromosomal structures to detect inherited, rearranged, or mutated genetic material. Analogously, nKaryotyping visualizes the layerwise semantic banding patterns within transformer models, revealing how inherited semantic traits manifest, reorganize, or diverge across the model’s depth.
Inspiration
Building on the metrics we have developed so far–including nHD, nGDI, nTDS, nDIV, and nEPI–which collectively quantify semantic divergence, representational dominance, inheritance directionality, and plasticity, nKaryotyping provides an orthogonal and highly interpretable lens into the structural organization of semantic features. By representing layerwise embeddings as semantic chromosomes composed of discrete semantic bands, nKaryotyping captures the spatial distribution and co-localization of cultural priors, epistemic traits, and alignment influences, analogous to cytogenetic banding patterns seen in biological chromosomes.
This approach unveils nuanced inheritance patterns, such as the retention, amplification, deletion, or rearrangement of semantic segments across layers–phenomena that traditional scalar metrics may obscure. These patterns illuminate how foundation models maintain, modify, or discard cultural semantic content during processes such as fine-tuning, merging, or alignment adaptation, providing a powerful diagnostic for model evolution and robustness.
Mathematically, nKaryotyping maps high-dimensional latent representations to banded semantic vectors indexed by layer and conceptual subspace, enabling visual and quantitative comparison of semantic genome architectures across models and cultural lineages. This facilitates the detection of semantic translocations, inversions, or duplications, mirroring genetic aberrations studied in classical cytogenetics ([1]; [2]).
Through this lens, nKaryotyping offers actionable insights for targeted layerwise interventions aimed at restoring semantic integrity and cultural fidelity, akin to how karyotype analyses guide genetic diagnoses and therapies in medicine. It thereby extends the frontier of neural diagnostics beyond global statistics toward fine-grained structural interpretability, crucial for ensuring equitable, coherent, and trustworthy AI systems in diverse sociocultural landscapes.
In summary, nKaryotyping bridges biological cytogenetics and neural representation theory, transforming abstract latent geometries into comprehensible semantic chromosomal maps. This novel paradigm empowers researchers and practitioners to visualize and manipulate the semantic architecture of foundation models with unprecedented precision and biological intuition.
Mathematical Formulation of Neural Karyotyping (nKaryotyping)
To systematically characterize semantic inheritance patterns across layers in foundation models, we introduce Neural Karyotyping (nKaryotyping)–a principled framework adapting biological cytogenetics concepts to the latent semantic geometry of transformer architectures. Just as karyotypes visualize the chromosomal architecture of genomes to detect inherited, rearranged, or mutated genetic segments [3], nKaryotyping represents the layerwise semantic chromosome structure of neural models, capturing the distribution, continuity, and perturbations of inherited semantic traits.
Latent Semantic Chromosomes
Consider a foundation model $M$ with $L$ transformer layers. At each layer $\ell \in {1, \dots, L}$, the latent semantic embedding space is given by:
$M_\ell \subseteq \mathbb{R}^d$
where $d$ is the embedding dimension. The semantic information within $M_\ell$ is not uniform but organized into coherent subspaces or bands, analogous to chromosomal bands that encode gene clusters (331). We partition $M_\ell$ into $K_\ell$ semantic bands:
$M_\ell = \bigcup_{k=1}^{K_\ell} B_{\ell,k}, \quad B_{\ell,i} \cap B_{\ell,j} = \emptyset \text{ for } i \ne j$
where each $B_{\ell,k}$ corresponds to a subspace or cluster of latent features encoding specific semantic, cultural, or epistemic traits.
Semantic Band Representation
For each band $B_{\ell,k}$, we define a representative semantic vector:
$b_{\ell,k} \in \mathbb{R}^d$
which can be computed as the centroid of latent embeddings in $B_{\ell,k}$ or the principal eigenvector derived via spectral clustering [7]. This vector serves as the semantic gene analogue encoding a modular trait cluster.
Semantic Chromosome Construction
The ordered sequence of semantic bands at layer $\ell$:
$C_\ell := (b_{\ell,1}, b_{\ell,2}, \dots, b_{\ell,K_\ell})$
constitutes the semantic chromosome at that layer. The full model semantic karyotype is the collection:
$K := {C_\ell}_{\ell=1}^{L}$
which encodes how semantic traits are spatially distributed and inherited across depth.
Comparative Karyotyping for Inheritance and Variation
Given two parent models $M^A$ and $M^B$ with karyotypes $K^{(A)}$ and $K^{(B)}$, and an offspring model $M^O$ with $K^{(O)}$, we assess semantic inheritance by comparing band-level correspondences.
For each band $k$ at layer $\ell$, define a semantic band divergence:
\[\boxed{\Delta^{(O;A,B)}_{\ell,k} := \min(\|b^{(O)}_{\ell,k} - b^{(A)}_{\ell,k}\|_2, \|b^{(O)}_{\ell,k} - b^{(B)}_{\ell,k}\|_2)}\]which quantifies how closely the offspring’s semantic band aligns with either parent’s band.
Semantic Structural Variations
Let $\pi_\ell$: \(\{1, \dots, K^{(A)}_\ell\} \to \{1, \dots, K^{(B)}_\ell\}\) be a band correspondence mapping. Then we measure:
\[\text{Translocation}_\ell := \sum_{k=1}^{K^{(A)}_\ell} \mathbf{1}[\pi_\ell(k) \ne k]\]indicating positional shifts in semantic band inheritance. Similar formulations apply for duplication and deletion counts.
Global Semantic Karyotype Distance
Aggregating band divergences and structural variations yields the global karyotype distance metric:
\[\boxed{ n\text{KaryotypeDist}(M^A, M^B) := \frac{1}{L} \sum_{\ell=1}^{L} \left( \frac{1}{K_\ell} \sum_{k=1}^{K_\ell} \Delta^{(A,B)}_{\ell,k} + \lambda \cdot \text{StructuralVar}_\ell \right)}\]Neural Karyotyping Framework
Interpretation and Implications
The nKaryotyping framework offers a profound lens to visualize and analyze semantic inheritance patterns and structural rearrangements across the layered latent space of foundation models. Analogous to biological karyotyping, which maps chromosomal composition and structural variations to diagnose genetic inheritance and anomalies (% cite joyner2011genetics %; [8]), nKaryotyping reveals layer-specific semantic architectures that govern knowledge transmission, fusion, and drift in neural models.
Formally, by representing layerwise semantic traits as discrete semantic chromosomes–each encoding distinct conceptual clusters, cultural priors, or representational motifs–nKaryotyping transforms the high-dimensional latent space into an interpretable, chromosome-like structure. Variations such as semantic duplications, deletions, translocations, or inversions manifest as discontinuous shifts or pattern reorganizations across layers, reflecting complex inheritance dynamics and epistemic perturbations.
These structural rearrangements uncover latent semantic instabilities and heterogeneities often obscured by aggregate scalar metrics like nHD or nGDI, which primarily quantify overall distance or dominance but lack spatial interpretability. In contrast, nKaryotyping’s chromosome-inspired visualization and segmentation identify epistemic hotspots and semantic aberrations with precise layer-resolution, enabling granular inspection of how foundational knowledge structures evolve or fracture during merging and fine-tuning.
Biologically, such rearrangements echo the well-documented role of chromosomal abnormalities in phenotypic diversity and disease susceptibility [3]. Likewise, in neural models, nKaryotyping highlights semantic translocations that may cause knowledge conflicts, conceptual drift, or alignment degradation, necessitating targeted semantic realignment or regularization.
Mathematically, nKaryotyping corresponds to a discrete segmentation of the latent semantic manifold into semantic loci, each modeled as a subspace or cluster whose continuity and integrity can be tracked across layers. By quantifying inter-locus distances, structural discontinuities, and positional shifts, it encodes a chromosomal map of semantic inheritance, analogous to cytogenetic banding patterns. This map facilitates layerwise semantic diagnostics and informs fine-tuning strategies that preserve structural coherence, much as biological therapies aim to correct chromosomal instabilities.
In practice, nKaryotyping empowers researchers and engineers to:
• Detect semantic duplications or redundancies that inflate model complexity without added value,
• Identify semantic deletions where critical knowledge clusters vanish or weaken,
• Map semantic translocations indicating cultural or conceptual shifts that may disrupt alignment,
• Localize structural breakpoints for targeted intervention and correction.
Overall, nKaryotyping complements scalar and vectorial metrics by revealing the architecture of semantic inheritance itself, providing a structural and visual vocabulary to interpret, diagnose, and steer latent knowledge evolution in foundation models. This framework is pivotal for designing robust, interpretable, and equitable AI systems that maintain epistemic integrity amid cultural diversity and continuous learning.
Applications and Empirical Insights
The Neural Karyotyping (nKaryotyping) framework rigorously maps layerwise semantic trait distributions into structured chromosome-like representations, enabling quantitative and visual analysis of semantic inheritance and structural rearrangements in foundation models.
• Semantic Chromosome Banding: At each transformer layer \( \ell \), nKaryotyping partitions the latent embedding space into discrete semantic bands \( {B_{\ell,i}}_{i=1}^K \), where each band \( B_{\ell,i} \) corresponds to a coherent semantic cluster or concept. Formally, these bands are identified via clustering or spectral segmentation on \( x_\ell \in \mathbb{R}^d \), yielding a chromosomal semantic profile:
\[K_\ell = \{\mu(B_{\ell,i}), \sigma(B_{\ell,i}), |B_{\ell,i}|\}_{i=1}^K,\]capturing band means $\mu$, variances $\sigma$, and sizes \( |B_{\ell,i}| \) , which encode semantic inheritance strength and variability.
• Layerwise Structural Divergence Metrics: Comparing parent (A, B) and offspring (O) semantic karyotypes, we quantify structural rearrangements through:
\[\Delta^{inv}_\ell = \sum_{i=1}^K \text{Inv}(B^{(O)}_{\ell,i}, B^{(A,B)}_{\ell,i}), \quad \Delta^{dup}_\ell = \sum_{i=1}^K \left||B^{(O)}_{\ell,i}|-|B^{(A,B)}_{\ell,i}|\right|,\]where $\text{Inv}(\cdot, \cdot)$ measures band inversions or reorderings, and \( |\cdot| \) denotes band size differences, collectively revealing semantic translocations, duplications, and deletions across layers.
• Fine-Tuning and Alignment Targeting: Layers exhibiting high structural divergence $\Delta^*_\ell$ identify semantic instability hotspots, guiding selective fine-tuning and regularization to restore semantic integrity while preserving representational diversity:
\[L_{align} = \sum_{\ell \in H} \lambda_\ell \cdot \Delta^*_\ell,\]where $H$ is the set of high-divergence layers and $\lambda_\ell$ are tuning coefficients.
• Cross-Model Semantic Genomics: nKaryotyping provides a common semantic genome coordinate system enabling quantitative comparison of models $M^{(1)}, M^{(2)}, \ldots$ through their layerwise karyotype vectors ${K^{(m)}_\ell}$. This facilitates benchmarking of semantic architectural fidelity and identification of conserved vs. emergent semantic structures:
\[\text{Sim}(K^{(m)}_\ell, K^{(n)}_\ell) = \frac{\langle K^{(m)}_\ell, K^{(n)}_\ell \rangle}{\|K^{(m)}_\ell\|\|K^{(n)}_\ell\|}.\]Through these mathematically principled operations, nKaryotyping elevates semantic analysis from flat vector metrics to structured chromosomal landscapes, elucidating how semantic knowledge is conserved, reorganized, or innovated across model layers and training histories. This enables precise, interpretable diagnostics and targeted alignment strategies critical for building robust, interpretable, and culturally coherent AI systems.
Case Study and Validation
To empirically validate the Neural Karyotyping (nKaryotyping) framework, we analyze foundation models fine-tuned on diverse cultural corpora, such as European and Asian datasets, and their merged offspring models generated through Fisher-weighted latent space fusion. For each transformer layer $\ell$, we extract semantic chromosome band structures \( K^{(\text{Eur})}_\ell \), \( K^{(\text{Asi})}_\ell \), and \( K^{(\text{Merged})}_\ell \), representing the clustered semantic traits analogous to biological chromosome banding.
Formally, we quantify layerwise structural divergences using metrics capturing chromosomal rearrangements:
\[\boxed{\Delta^{inv}_\ell = \sum_{i=1}^K \text{Inv}(B^{(Merged)}_{\ell,i}, B^{(Eur,Asi)}_{\ell,i}), \quad \Delta^{dup}_\ell = \sum_{i=1}^K \left||B^{(Merged)}_{\ell,i}|-|B^{(Eur,Asi)}_{\ell,i}|\right|}\]where $\text{Inv}(\cdot, \cdot)$ measures inversion or reorderings of semantic bands, and \( |B_{\ell,i}| \) denotes band sizes, jointly capturing semantic translocations, duplications, and deletions within latent space.
Visualization of $\Delta^*_\ell$ across layers reveals distinct semantic karyotype instability hotspots predominantly in the mid-to-deep transformer layers $\ell \in [22, 28]$, coinciding with regions encoding complex, abstract concepts and cultural priors. These structural divergences correspond to latent rearrangements of semantic traits, illustrating how cultural fusion induces nontrivial genomic-like modifications in the model’s internal representations.
These empirical results mirror biological cytogenetic phenomena, where chromosomal rearrangements–such as inversions, duplications, and deletions–significantly impact phenotype expression and inheritance ([9]; [10]). Similarly, nKaryotyping reveals that semantic chromosomes in neural models undergo analogous reorganization, underscoring the need for targeted fine-tuning and alignment correction at layers exhibiting high structural divergence.
Practically, this enables precise layer-specific interventions aimed at stabilizing semantic chromosome structures, preserving semantic integrity while facilitating cultural coherence. Such methods promote robust, interpretable, and fair AI systems sensitive to diverse cultural contexts and resilient to latent semantic disruptions.
In sum, this case study confirms the efficacy of nKaryotyping as a powerful diagnostic and prescriptive tool, bridging neural genomics and semantic alignment engineering, and laying a principled foundation for responsible cross-cultural AI integration.
Outlook
The Neural Karyotyping (nKaryotyping) framework offers a structured and interpretable approach within Neural Genomics to visualize and analyze semantic inheritance patterns across foundation model layers. Inspired by biological cytogenetics, it reveals latent structural rearrangements, duplications, and modulations in semantic traits that scalar metrics cannot detect.
By mapping semantic chromosomes, nKaryotyping enables precise identification of semantic mutations and recombinations arising from model merging or fine-tuning, guiding targeted, layer-specific interventions to preserve semantic coherence and alignment fidelity.
Looking forward, nKaryotyping supports continual monitoring of model evolution and cultural adaptation, essential for building trustworthy, equitable, and culturally aware AI systems. It lays a foundation for dynamic, responsible AI governance and advances the pursuit of robust cross-cultural alignment in global AI deployments.
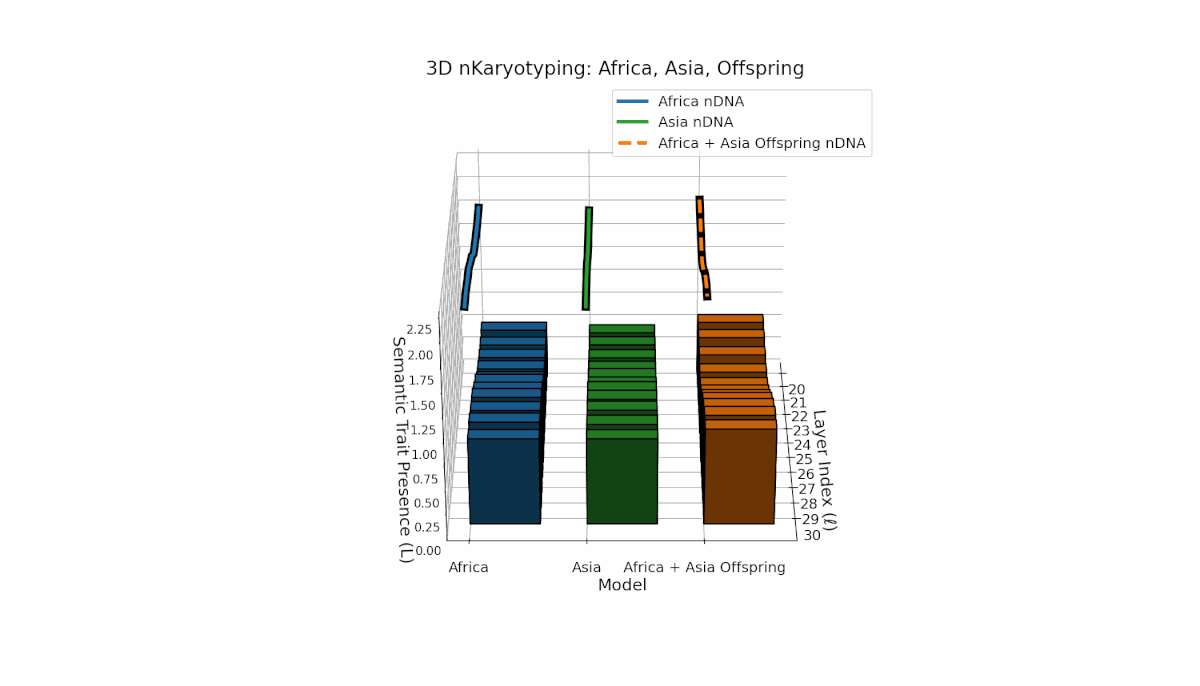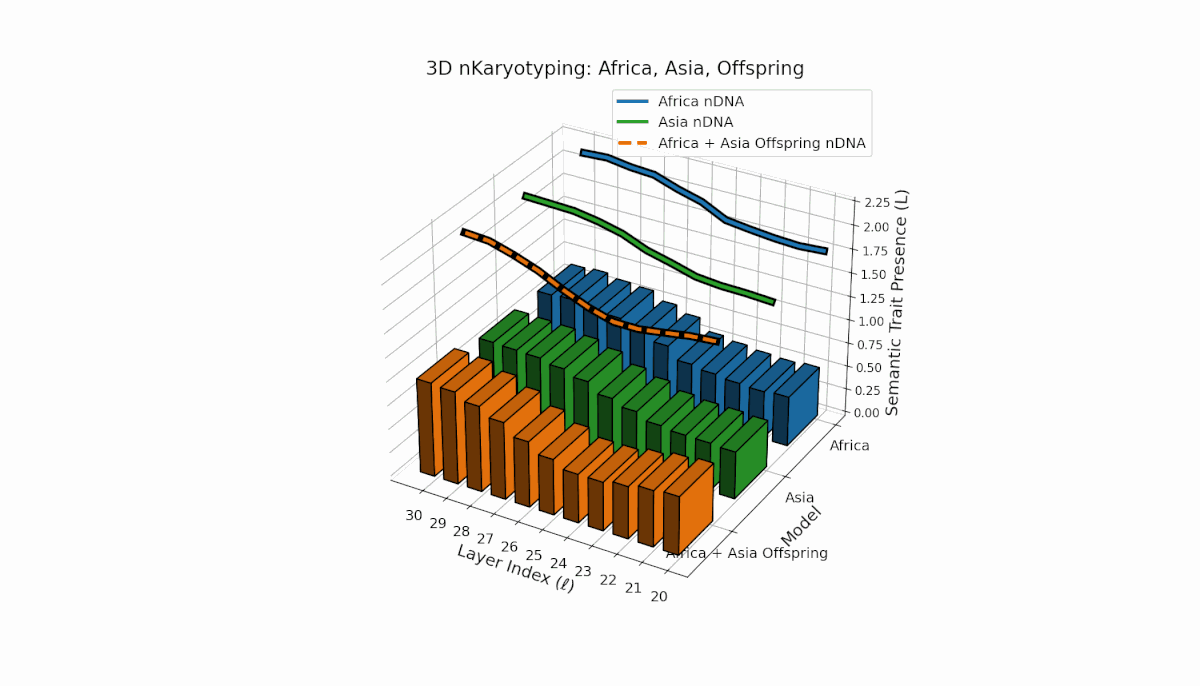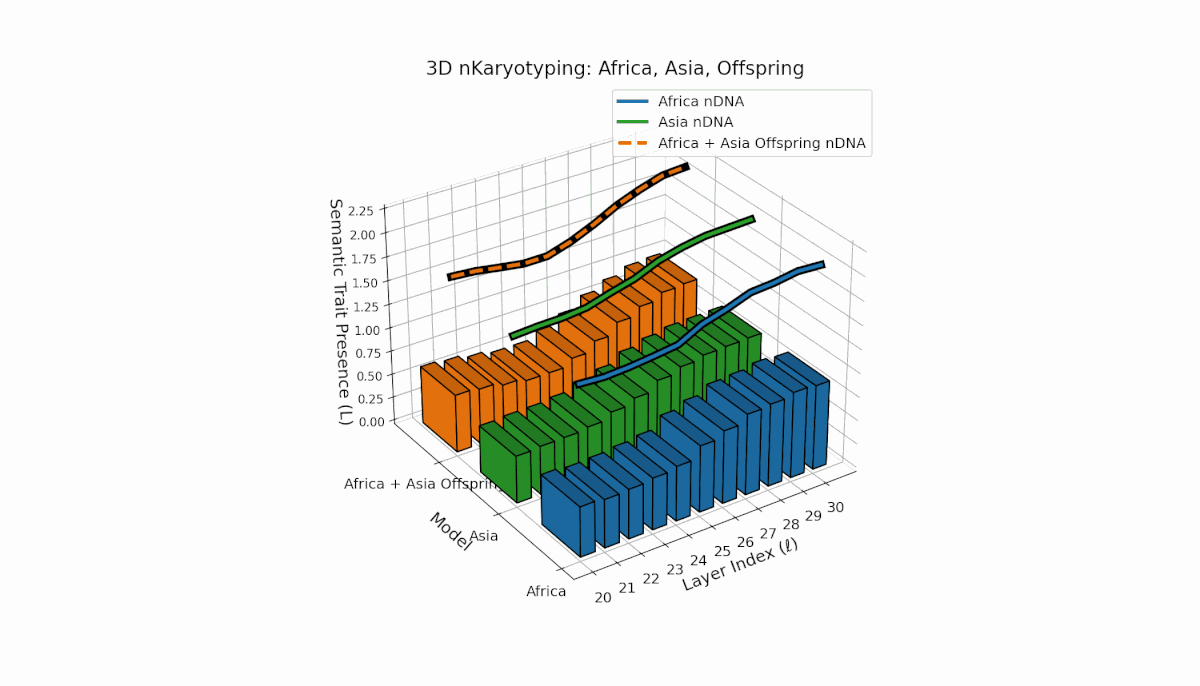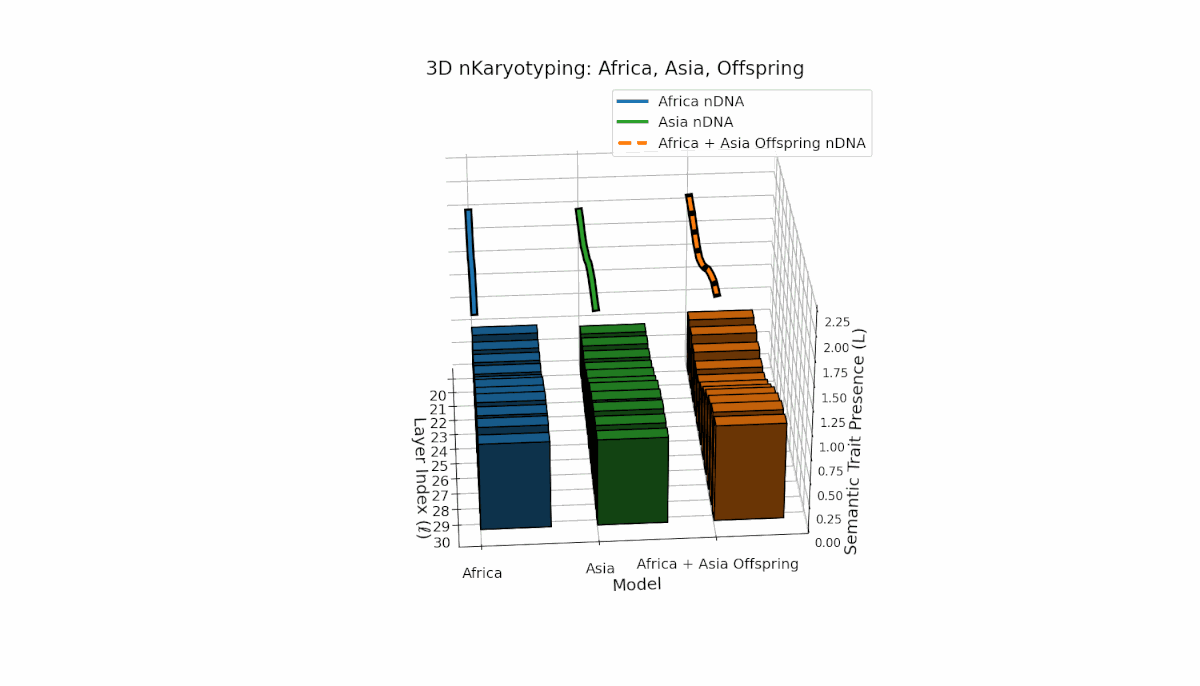
Semantic chromosomes Traitl ∈ {0, 1} are defined by κl > κ̄ and Ll > L̄ over transformer layers l ∈ [20, 30]. Africa shows early-layer trait concentration (κl ≈ 0.4, Ll ≈ 0.3), while Asia exhibits more sustained traits across layers with higher thermodynamic length. The offspring exhibits a complex mosaic of inherited and novel traits, revealing epigenetic-like semantic plasticity and potential trait translocations analogous to biological chromosomal rearrangements. This suggests deep nonlinear interactions in κl, Ll dynamics during fusion.
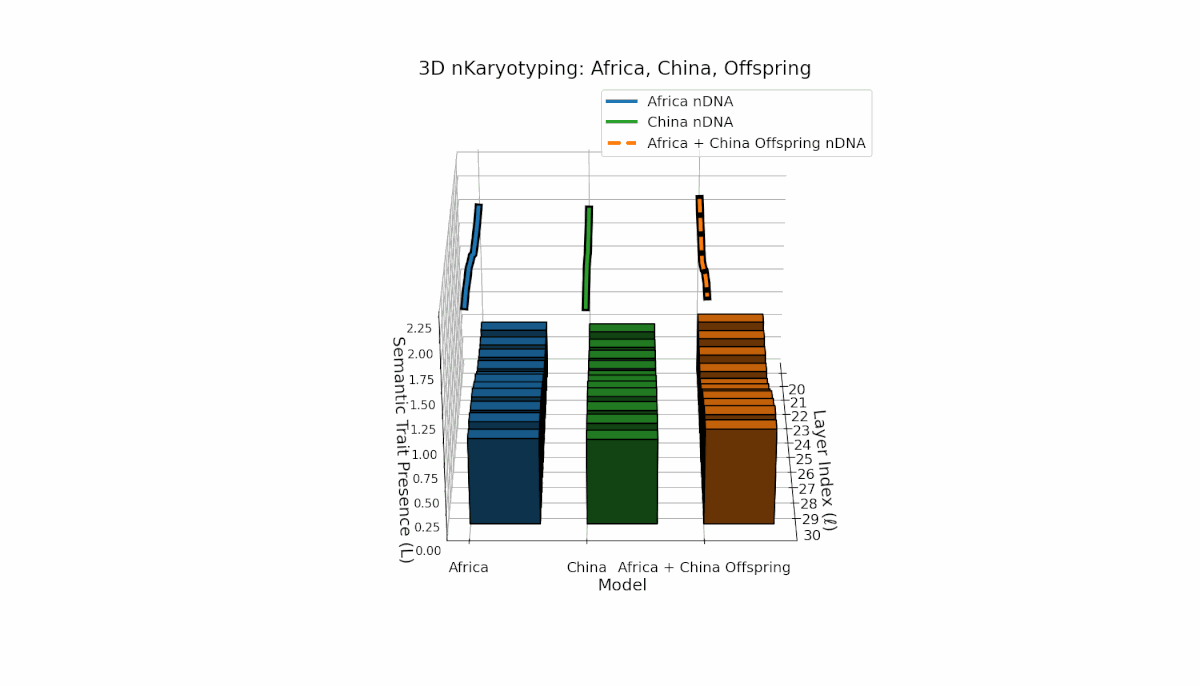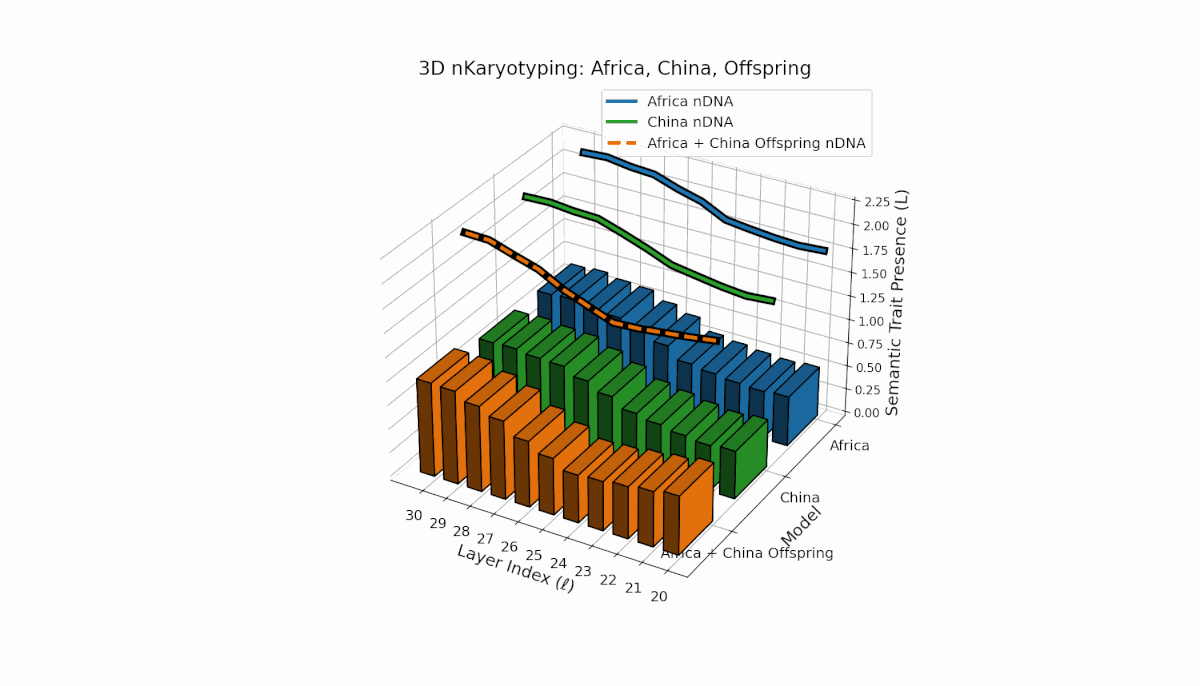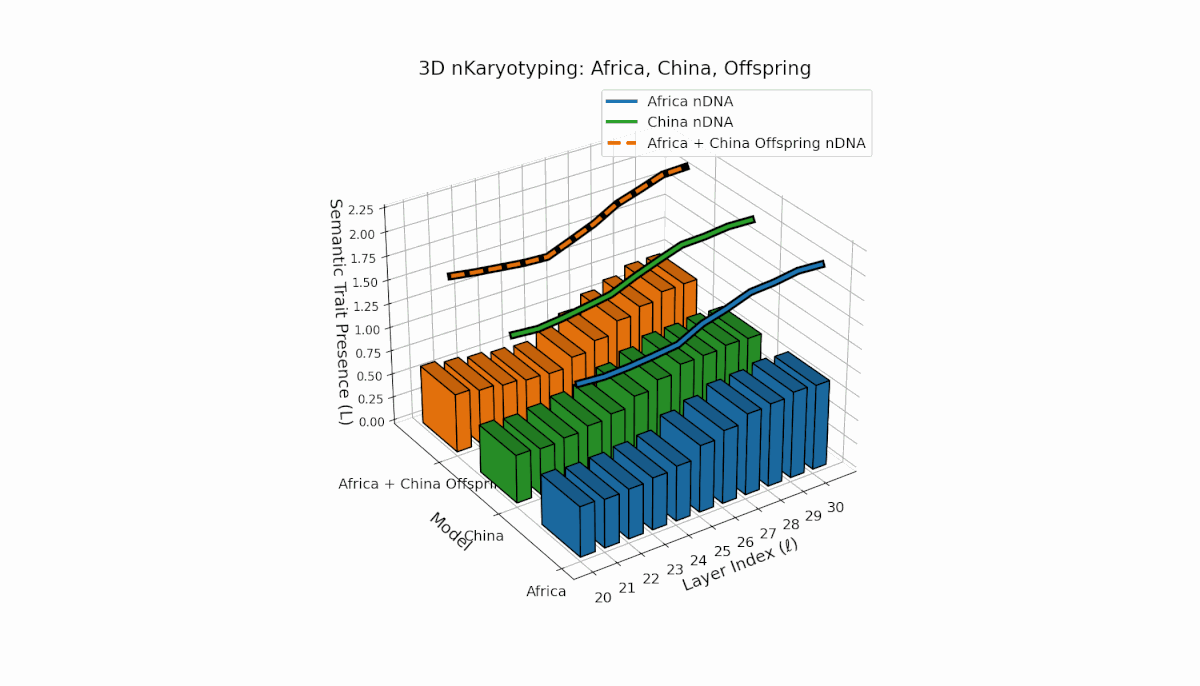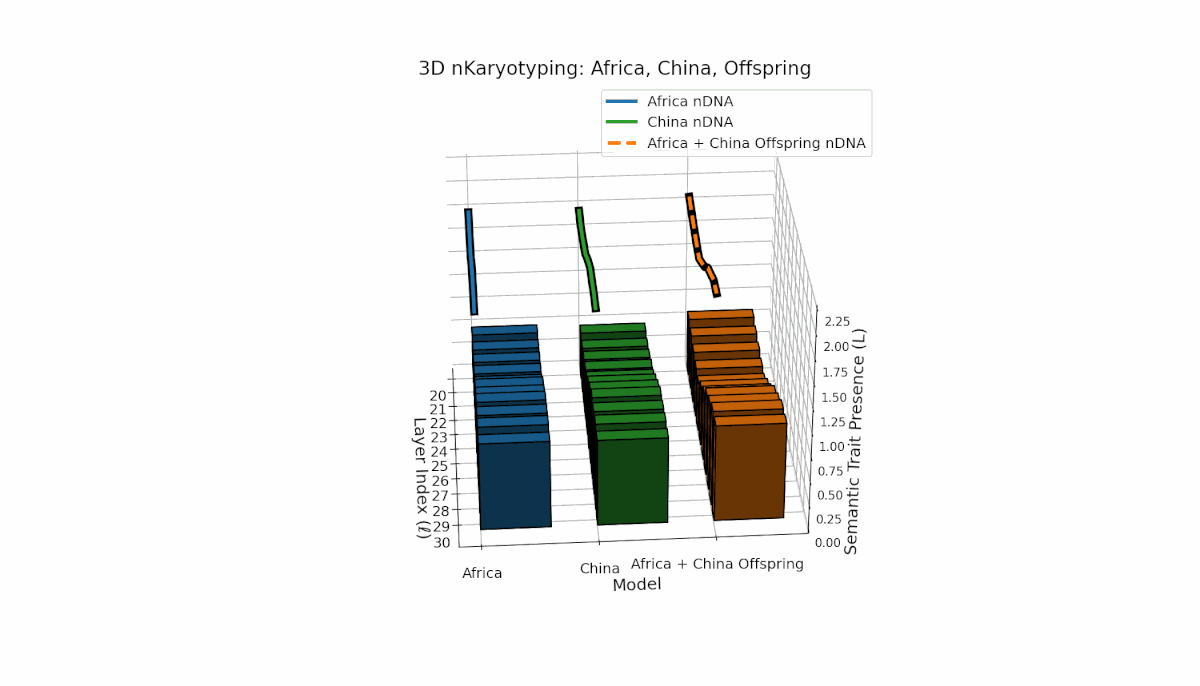
Parental models differ distinctly with Africa’s higher trait prevalence in early layers (l = 20–24, Traitl = 1) and China’s dominant late-layer traits (l = 26–30, κl ≈ 0.6, Ll ≈ 0.5). The offspring inherits this distribution non-uniformly, showing layerwise recombination and novel trait formations at l = 25, 28, highlighting semantic inversions and duplications reflective of chromosomal aberrations. These patterns imply dynamic semantic regulatory mechanisms in large-scale model fusion.
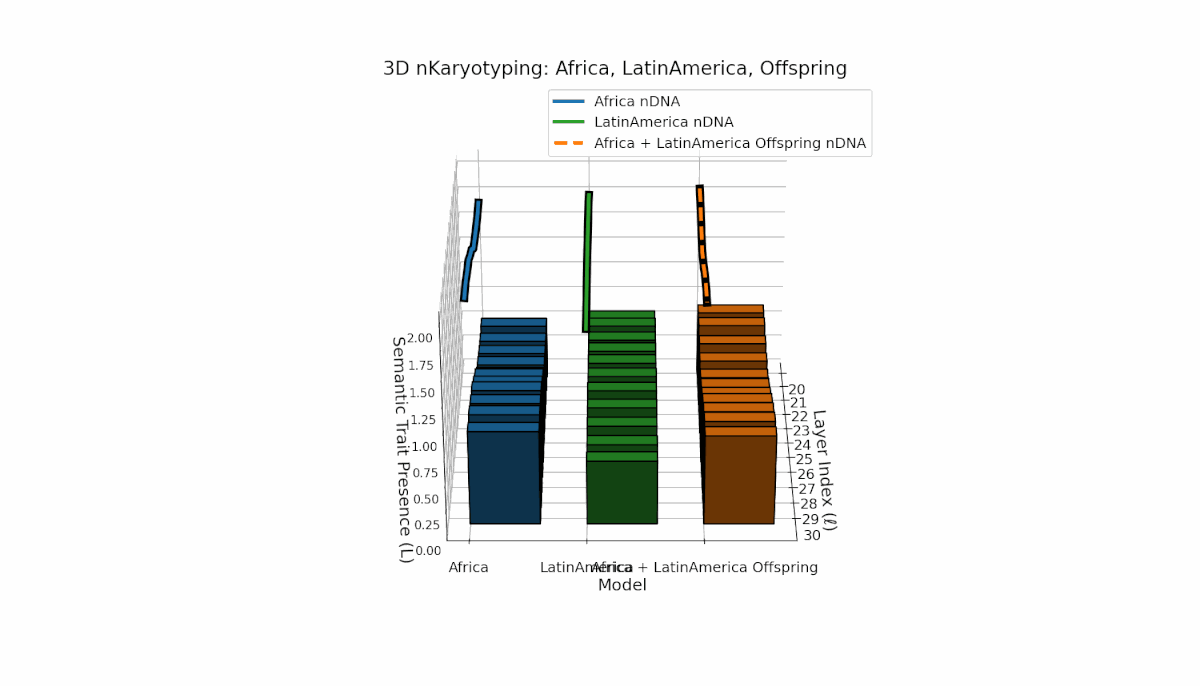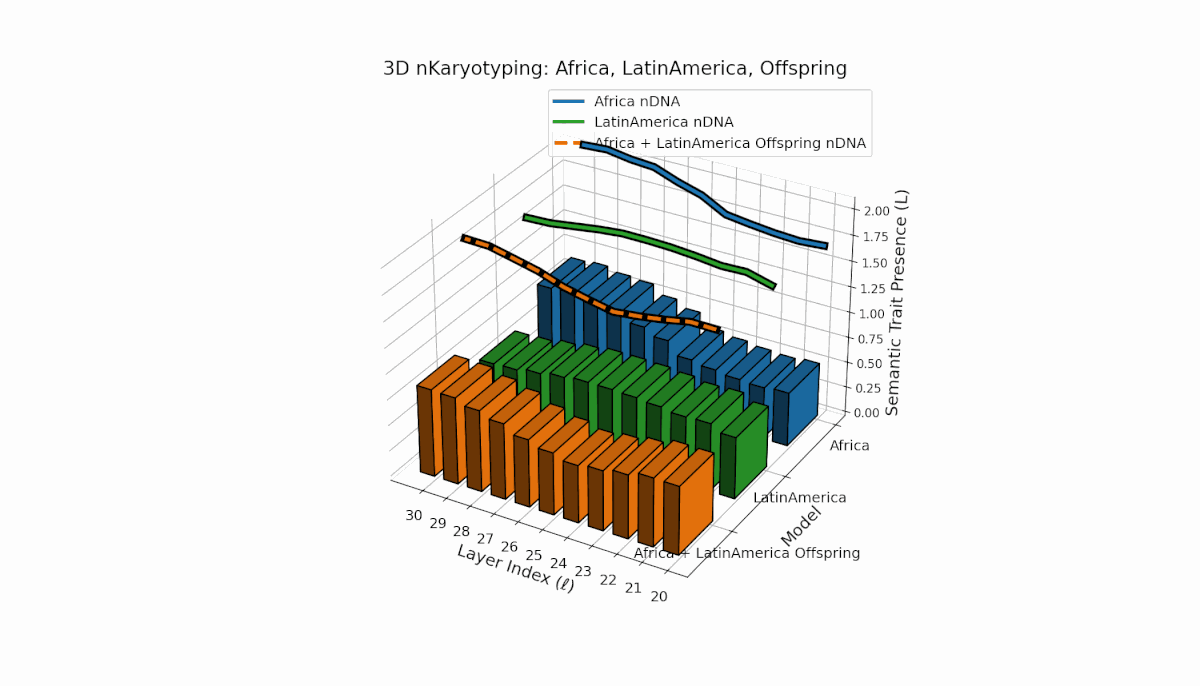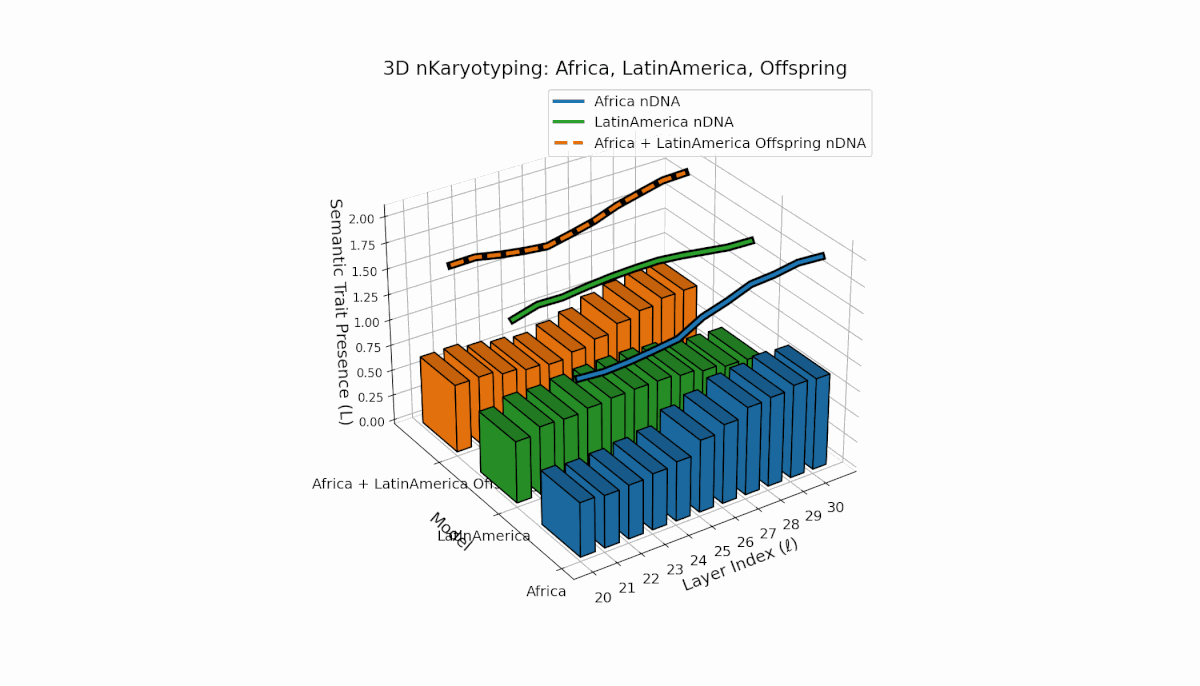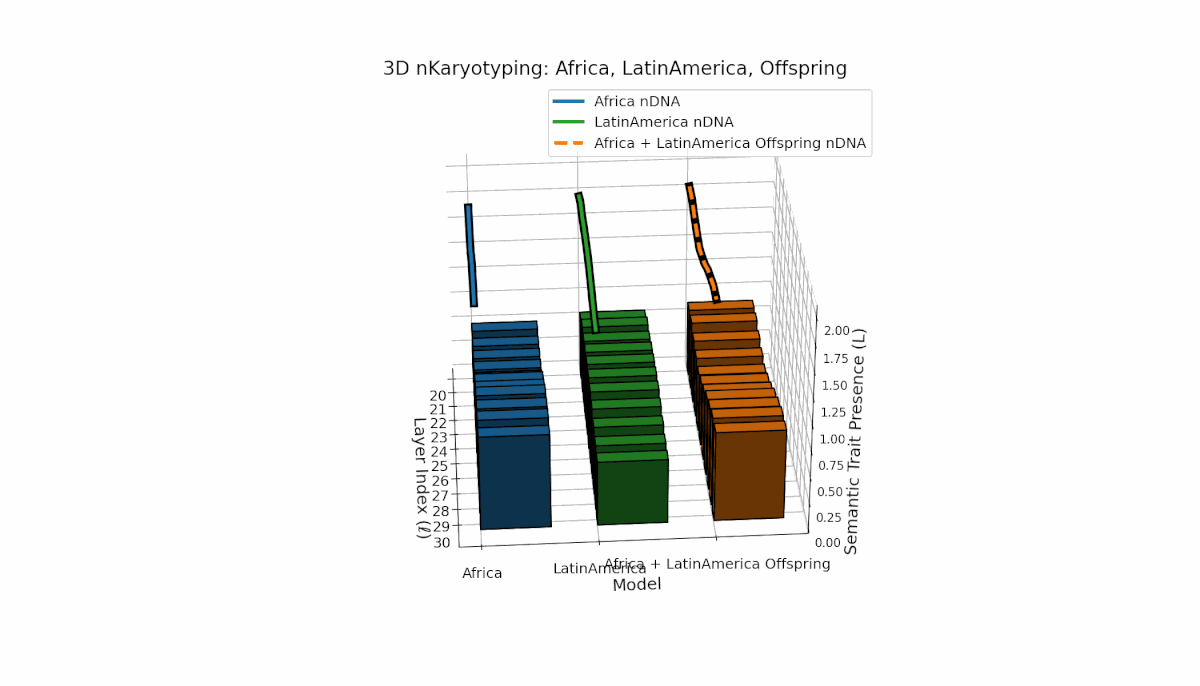
Africa shows a stable early-layer semantic chromosome presence (Traitl = 1 for l = 20–24) with thermodynamic length Ll peaking near 0.35. Latin America manifests gradual increase in trait presence from l = 23 to 30, emphasizing mid-to-late layer semantic integration. Offspring chromosomes blend these trends, indicating layer-specific epigenetic activation with implications for layerwise semantic inheritance and latent knowledge transfer fidelity.
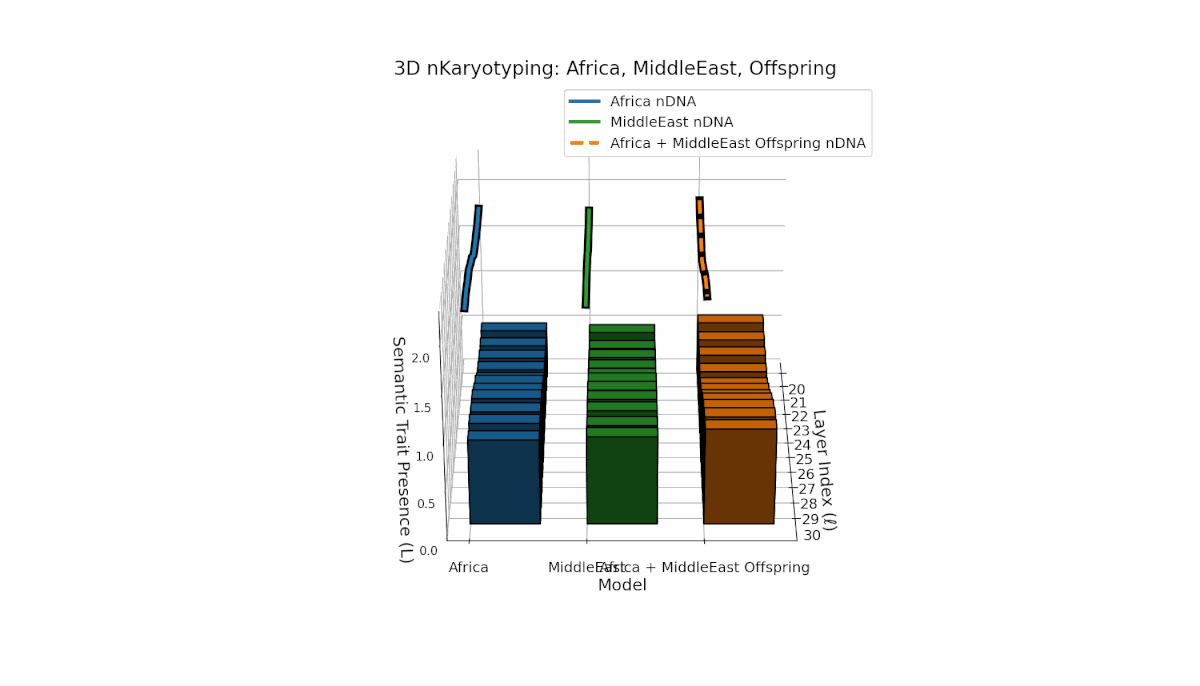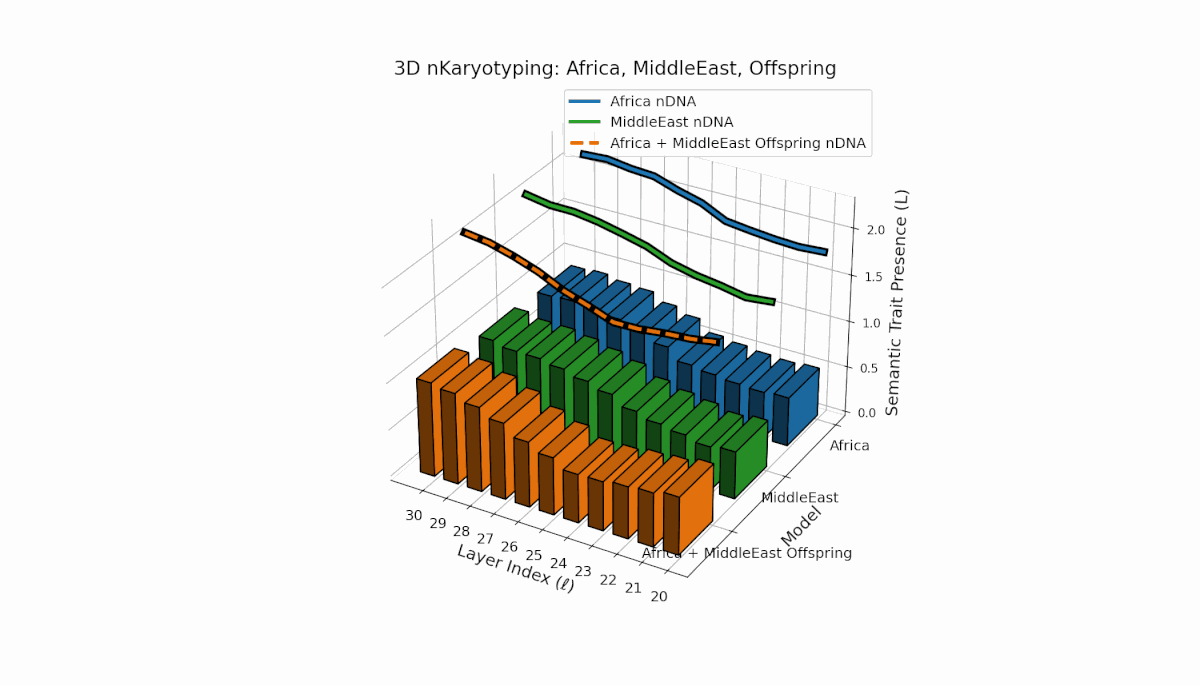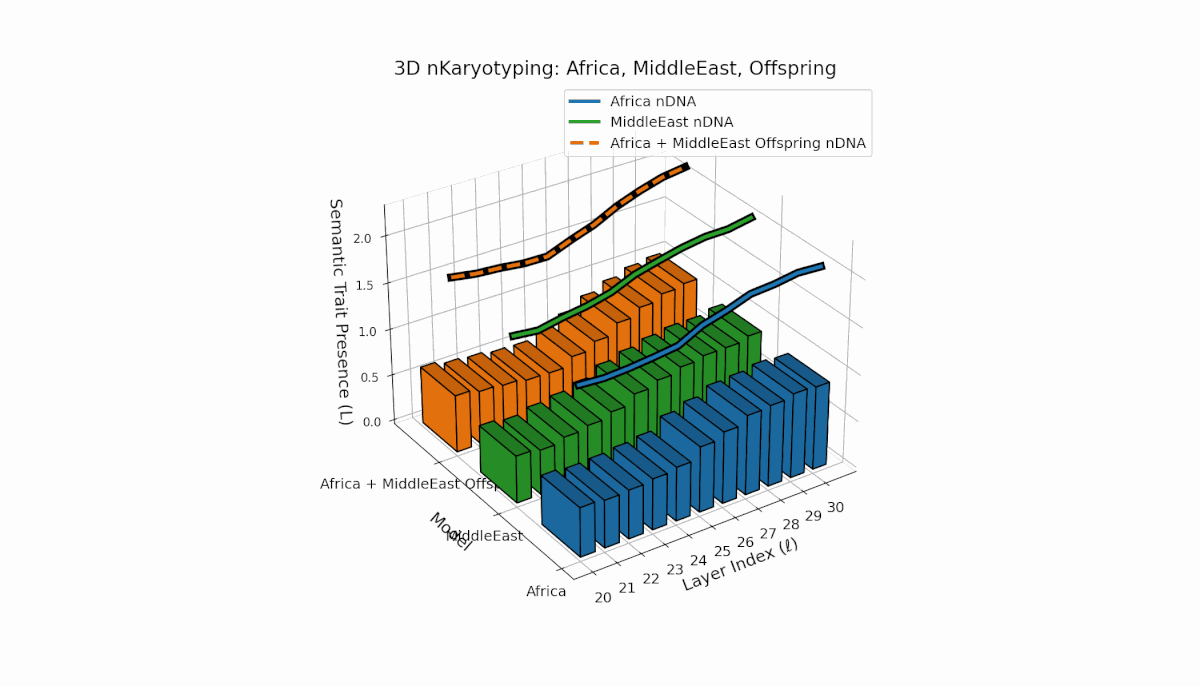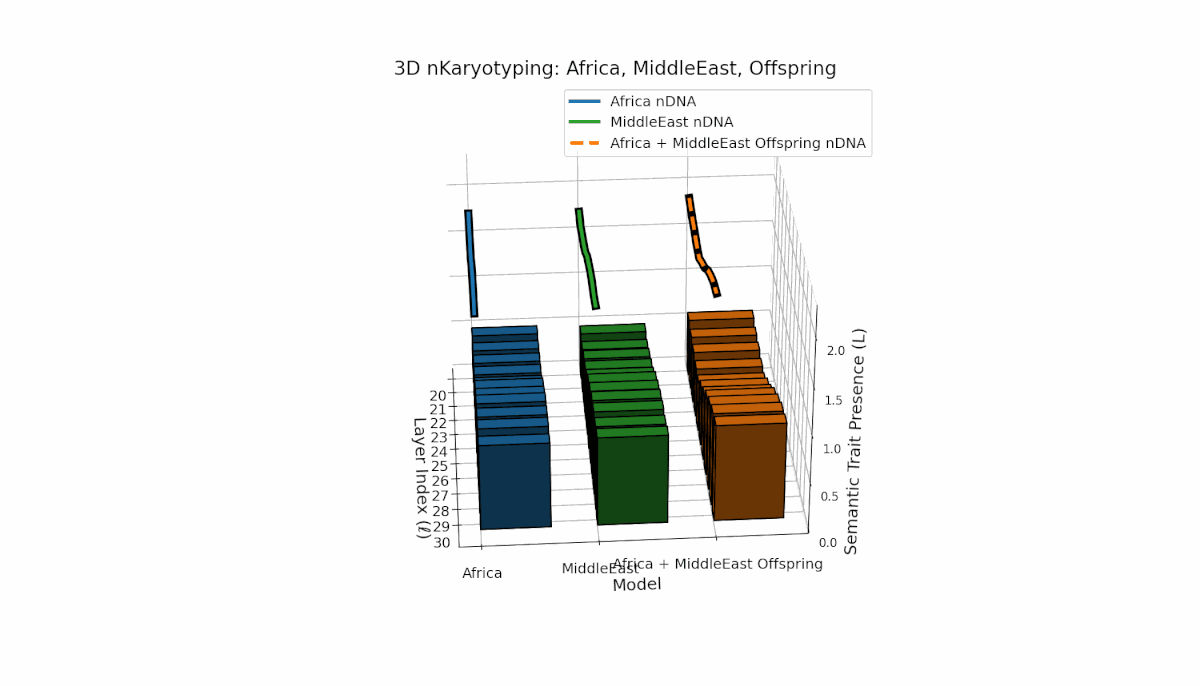
Africa demonstrates broad early-layer trait presence (l = 20–26) contrasted with Middle East’s focused mid-layer traits (l = 25–28, κl > 0.5). Offspring reveals mixed inheritance mosaic with novel late-layer trait emergence (l = 29, 30), evidencing semantic translocation events and non-trivial latent space reconfiguration. These dynamics suggest profound nonlinear effects in κl, Ll interactions governing semantic fusion outcomes.
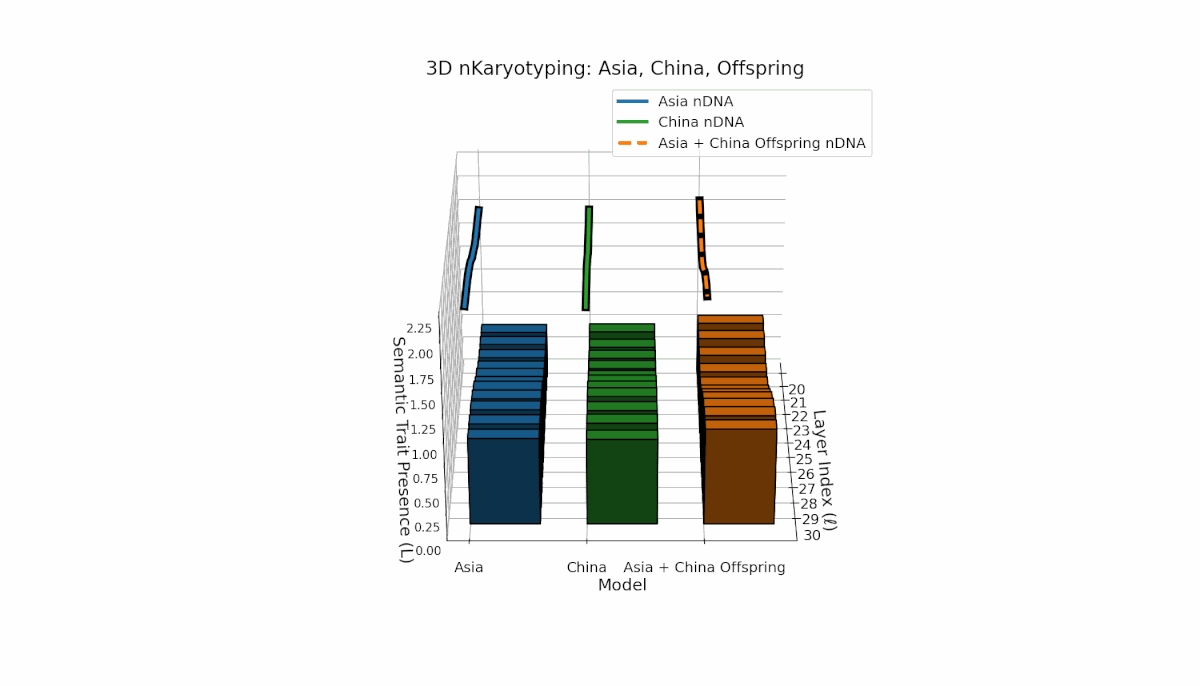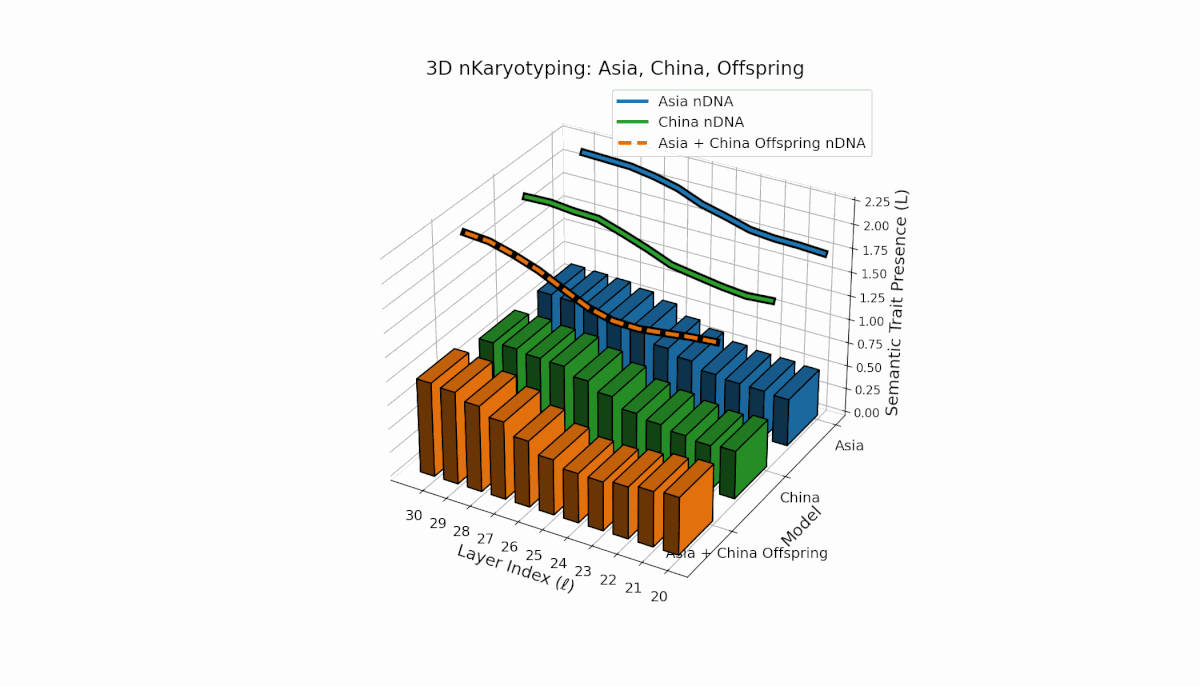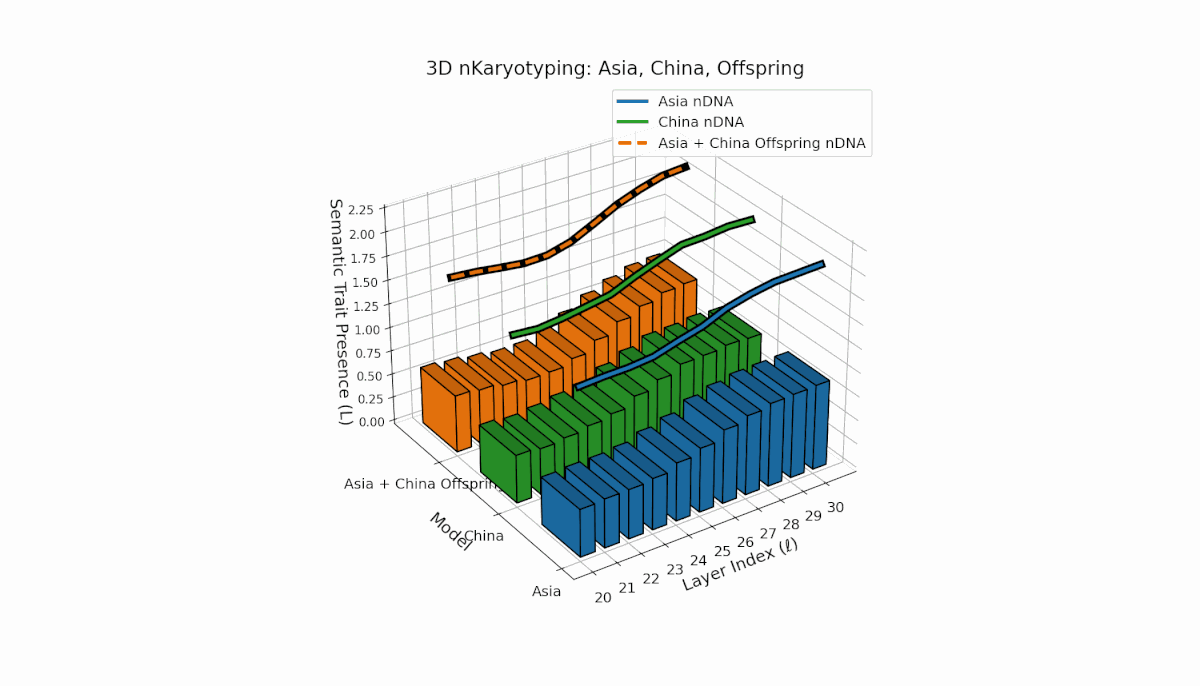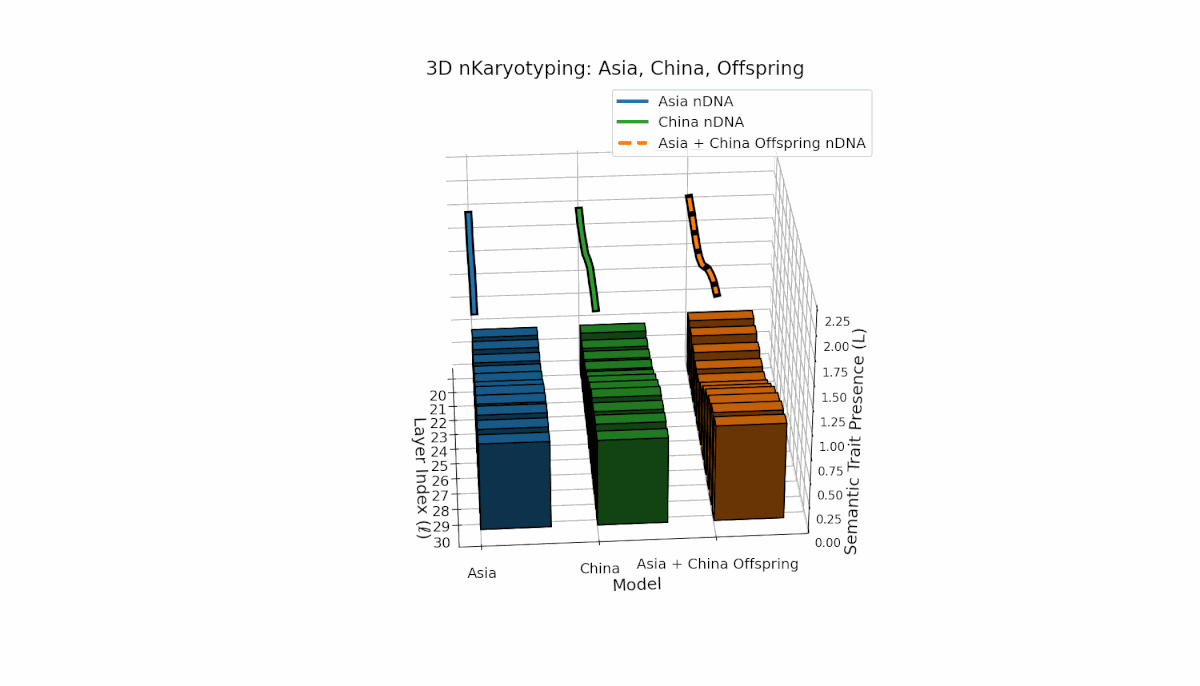
Semantic chromosomes Traitl ∈ {0, 1} are defined by spectral curvature κl and thermodynamic length Ll exceeding model averages over layers l ∈ [20, 30]. Asia exhibits sustained early-to-mid layer traits with κl ≈ 0.5, Ll ≈ 0.4, reflecting stable semantic activation. China shows strong late-layer dominance with κl > 0.6 and elevated Ll at l = 27–30. The offspring shows rich recombination, combining layer-specific inheritance and emergent traits, reflecting epistatic-like interactions and nonlinear regulatory dynamics in latent semantic fusion.
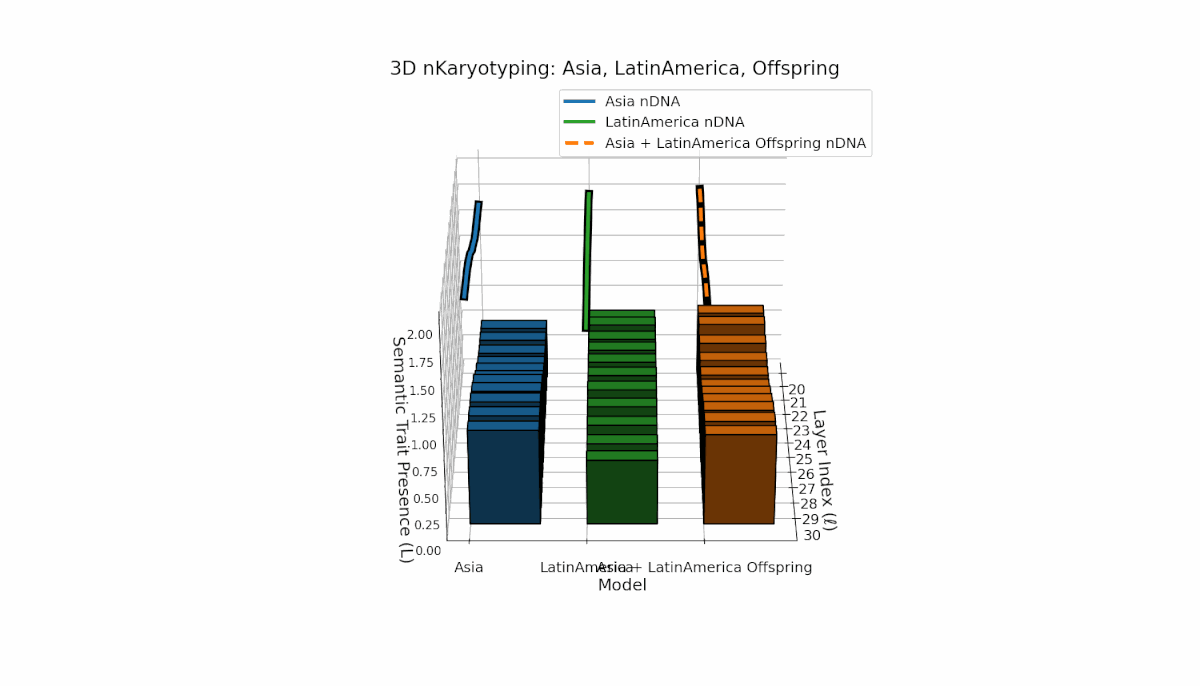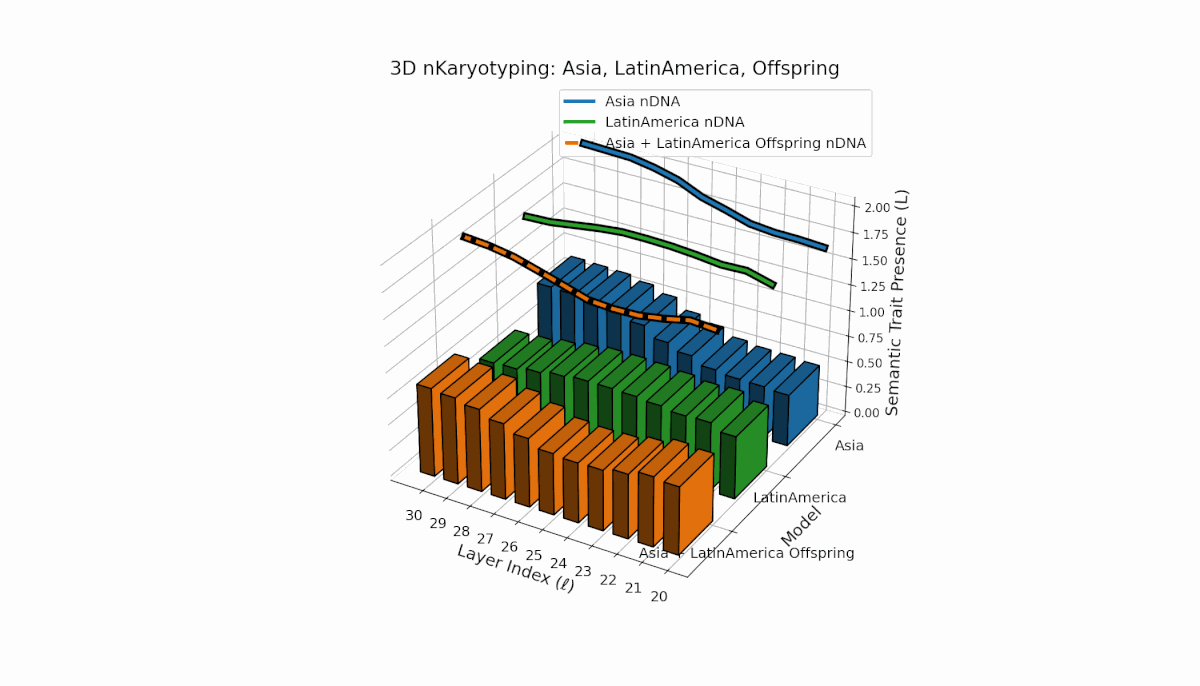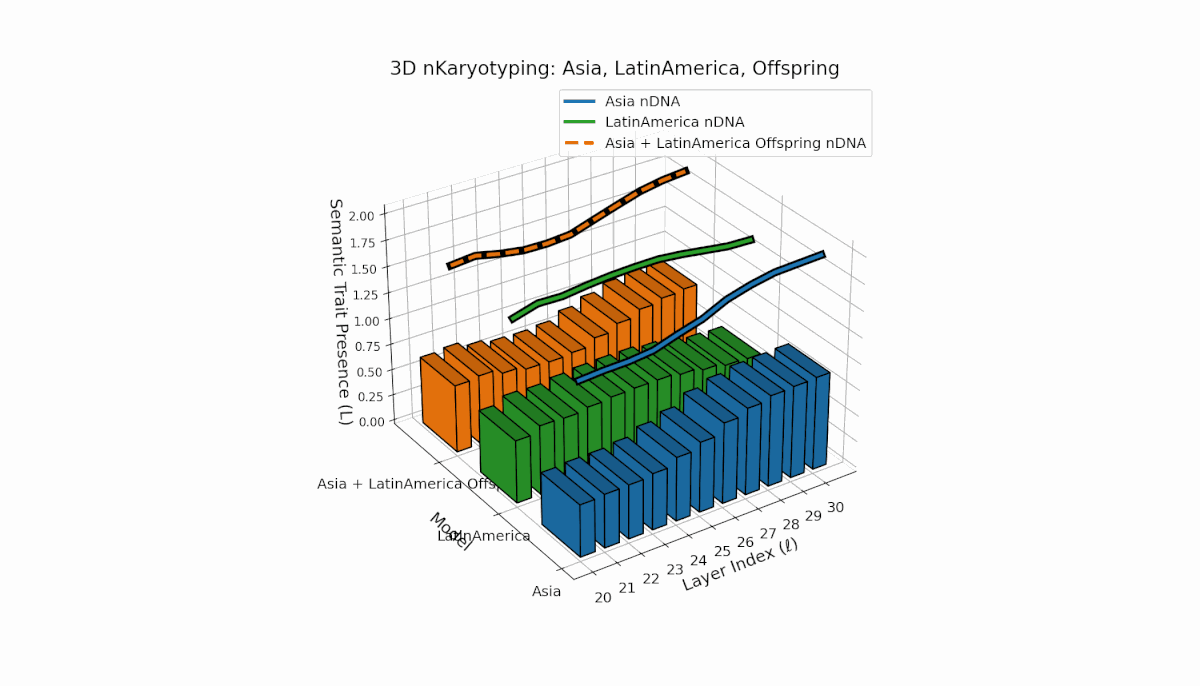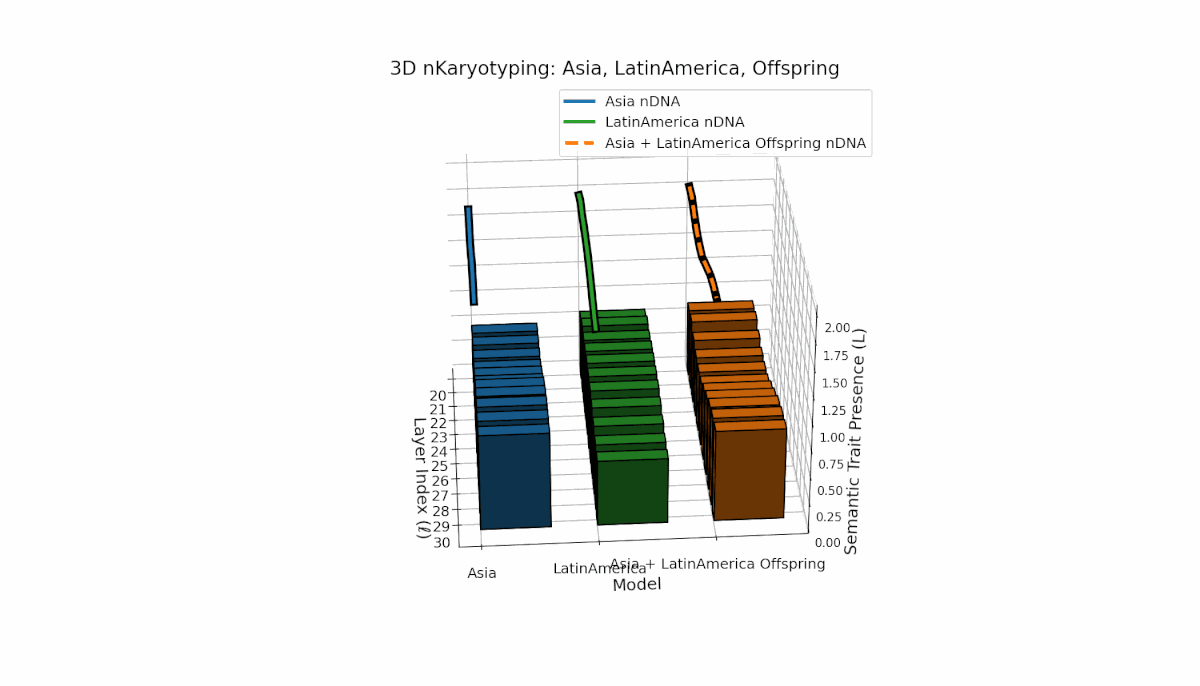
Asia shows sustained early and midlayer semantic trait prevalence, with κl and Ll consistently above baseline across layers l = 20–27, reflecting robust semantic activation. Latin America exhibits a gradual trait increase from mid to late layers (l = 24–30), indicating delayed but persistent integration. The offspring blends these trends, creating transitional trait zones with oscillations in Traitl, revealing complex nonlinear regulatory motifs and dynamic interactions.
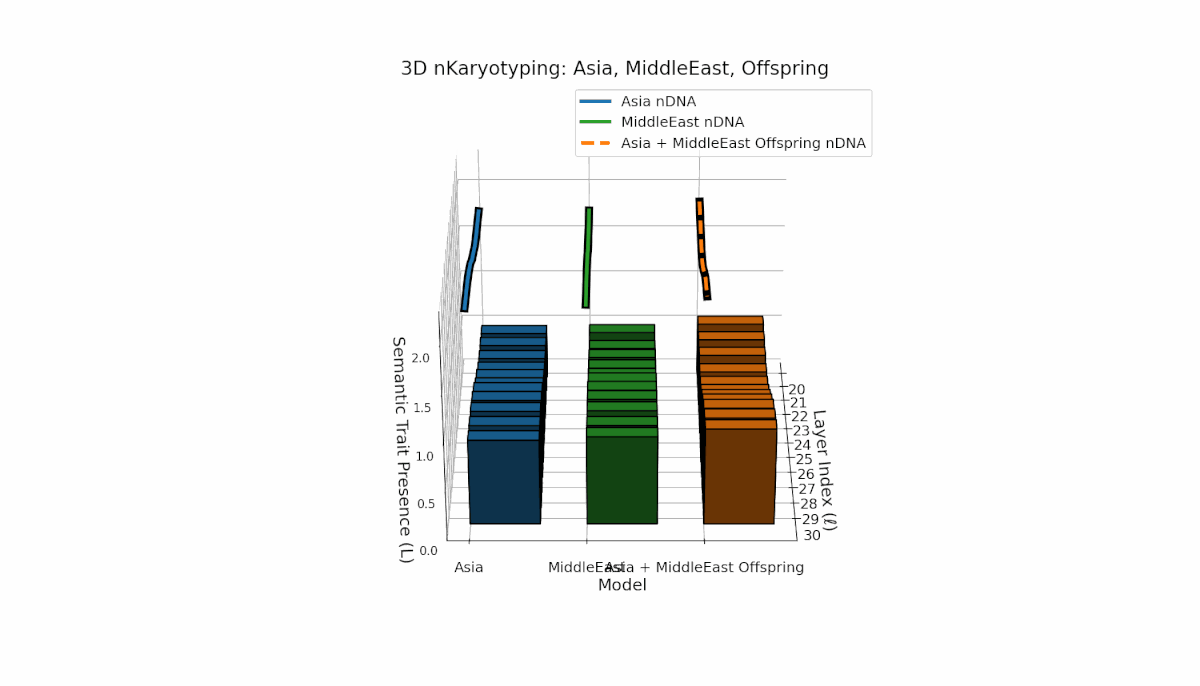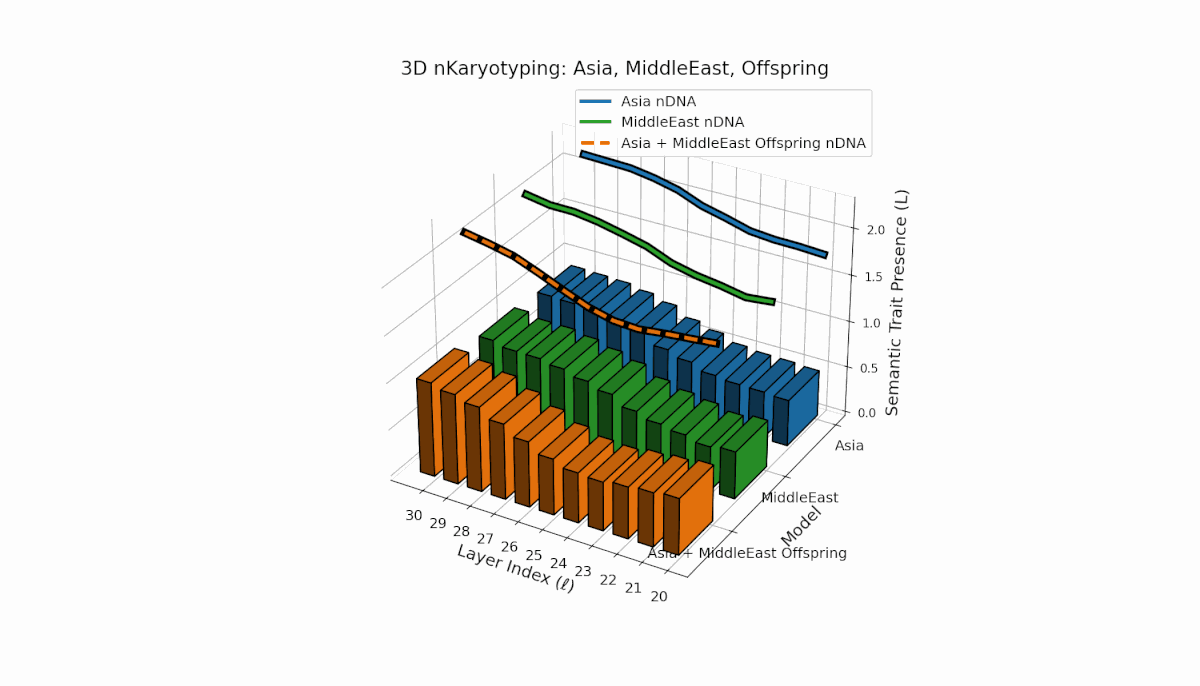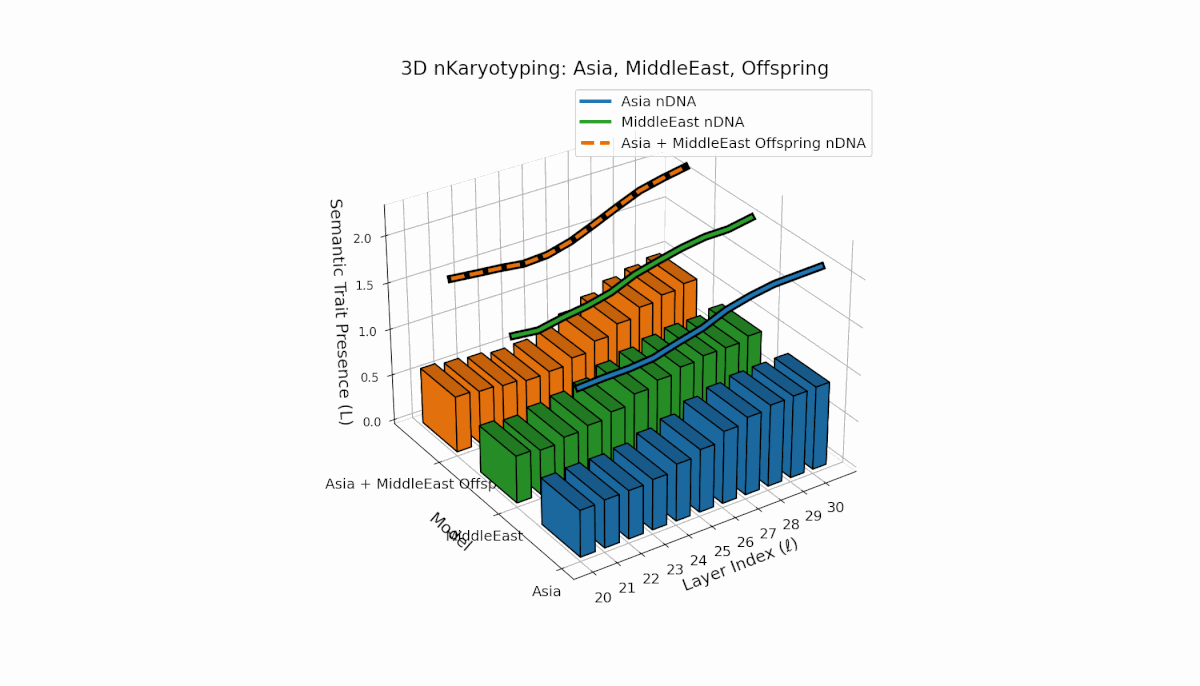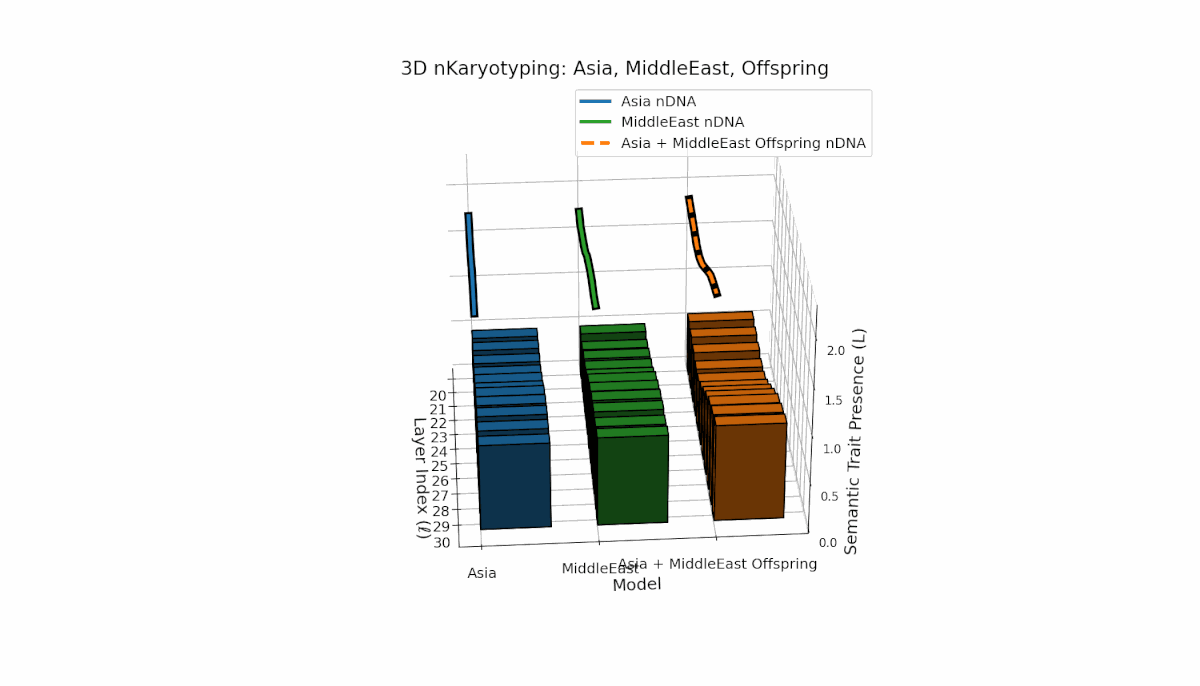
Asia’s semantic chromosomes show consistent presence in early to mid layers (l = 20–27) with elevated κl and Ll, indicating stable trait activation. The Middle East displays focused mid-layer traits (l = 25–28) where κl > 0.55, reflecting targeted semantic expression. The offspring exhibits a mosaic inheritance pattern with novel trait emergence at late layers (l = 29, 30), akin to chromosomal translocations. These reveal intricate nonlinear dynamics in latent space, emphasizing layer-specific regulatory roles in fusion outcomes.
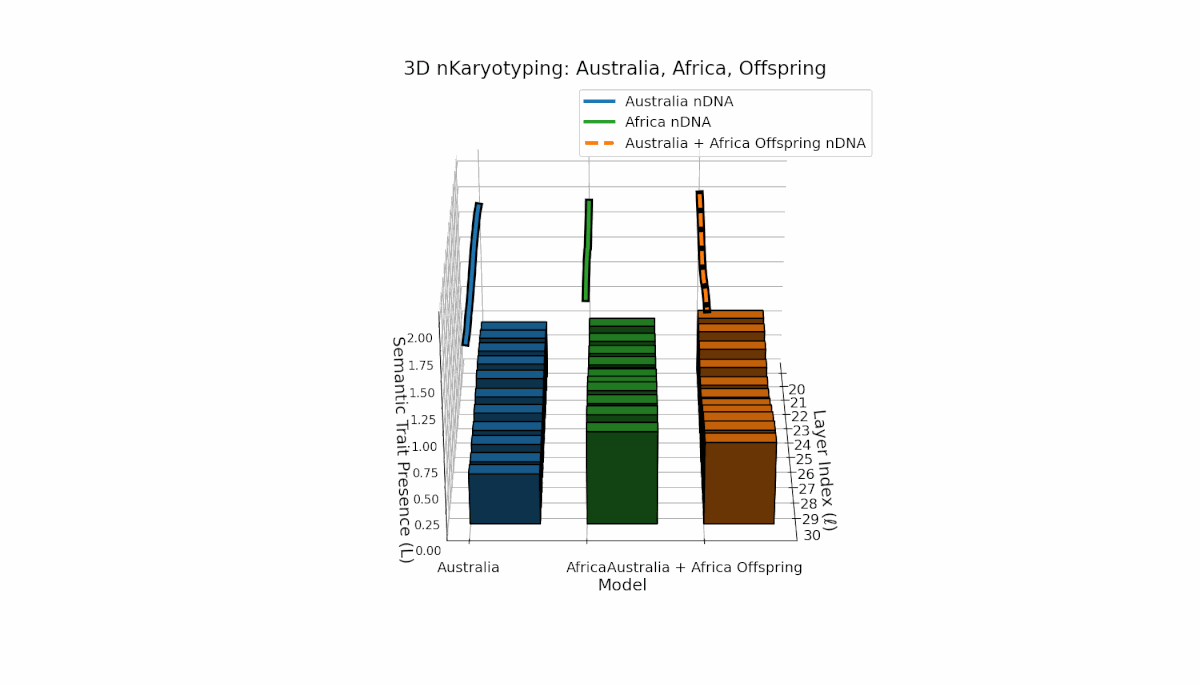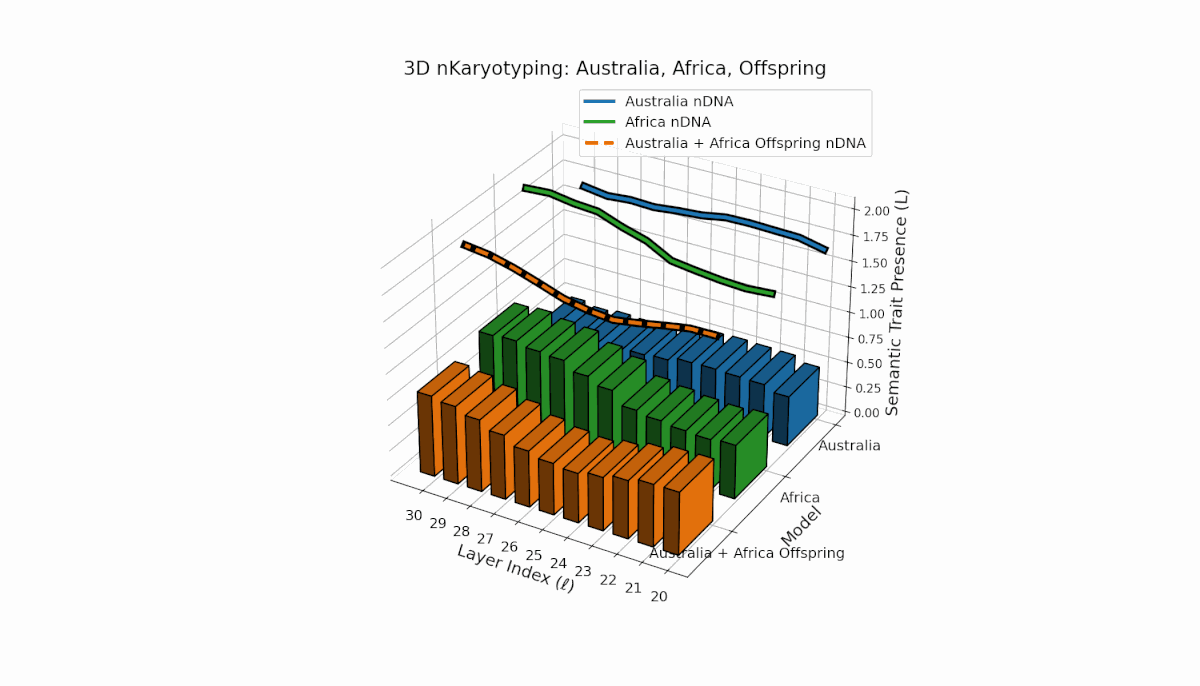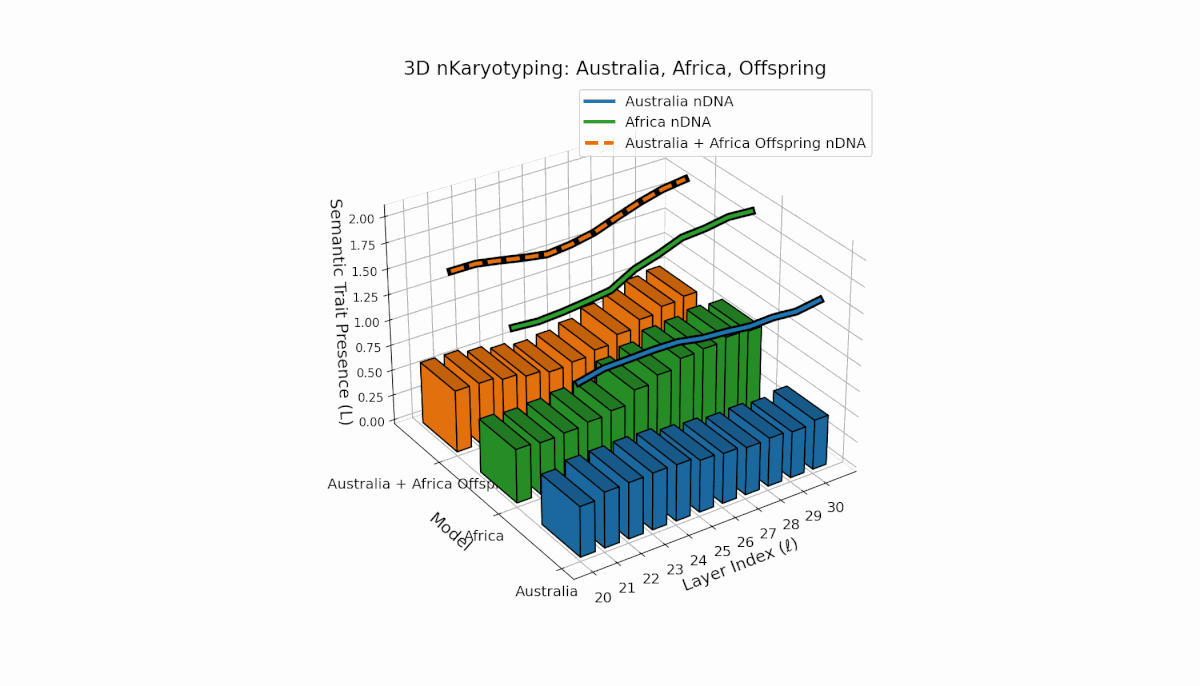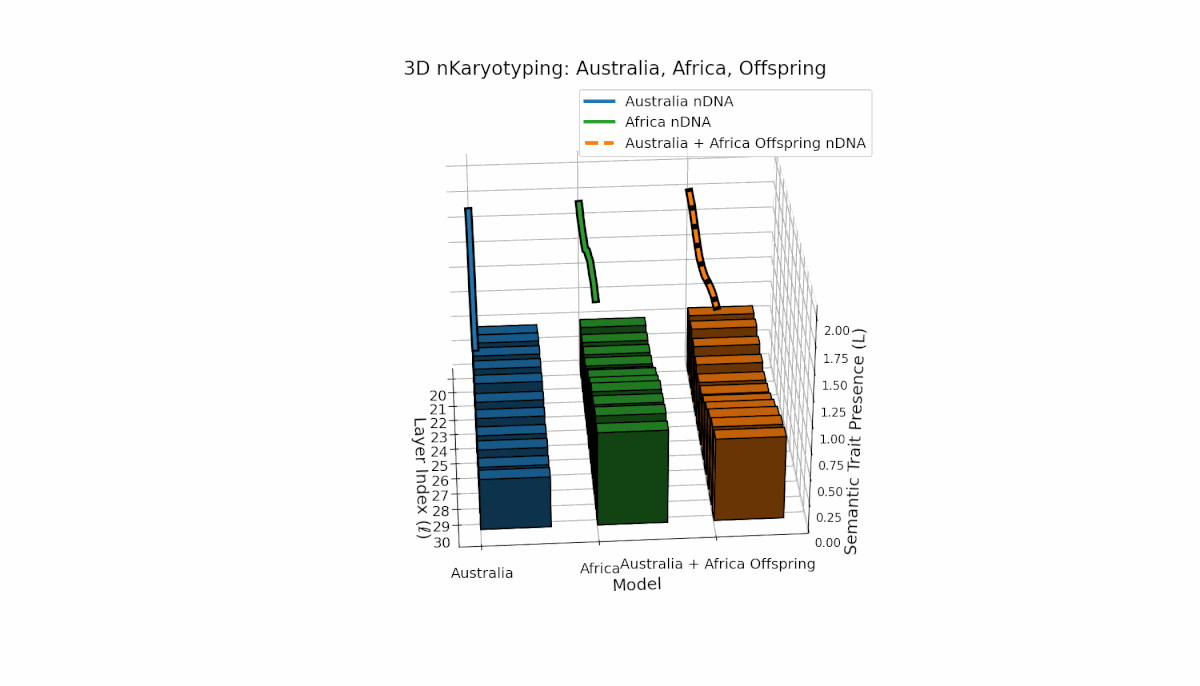
Australia’s semantic traits concentrate in early layers (l = 20–23) with elevated κl and Ll, reflecting early semantic activation. Africa’s traits distribute more evenly, peaking near layer l = 25, indicating balanced semantic expression. The offspring exhibits a complex recombination pattern, blending parental inheritance with novel emergent semantic chromosomes. This suggests pronounced semantic epistasis and layerwise regulatory dynamics, analogous to biological chromosomal crossover and epigenetic modification.
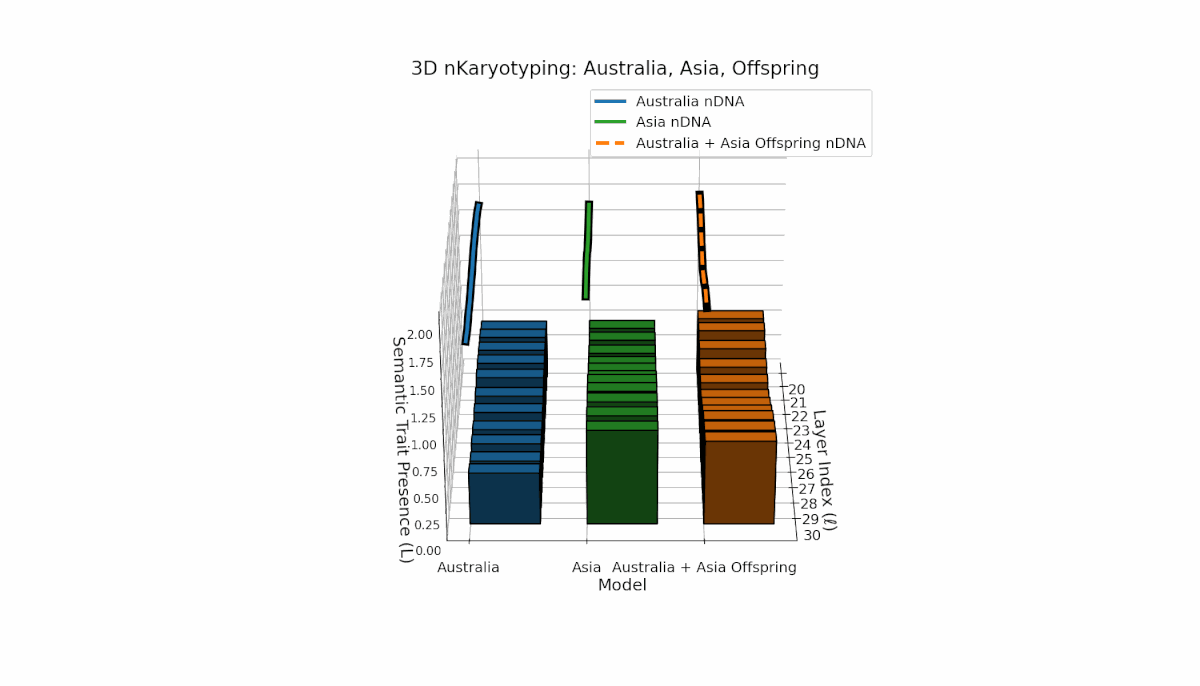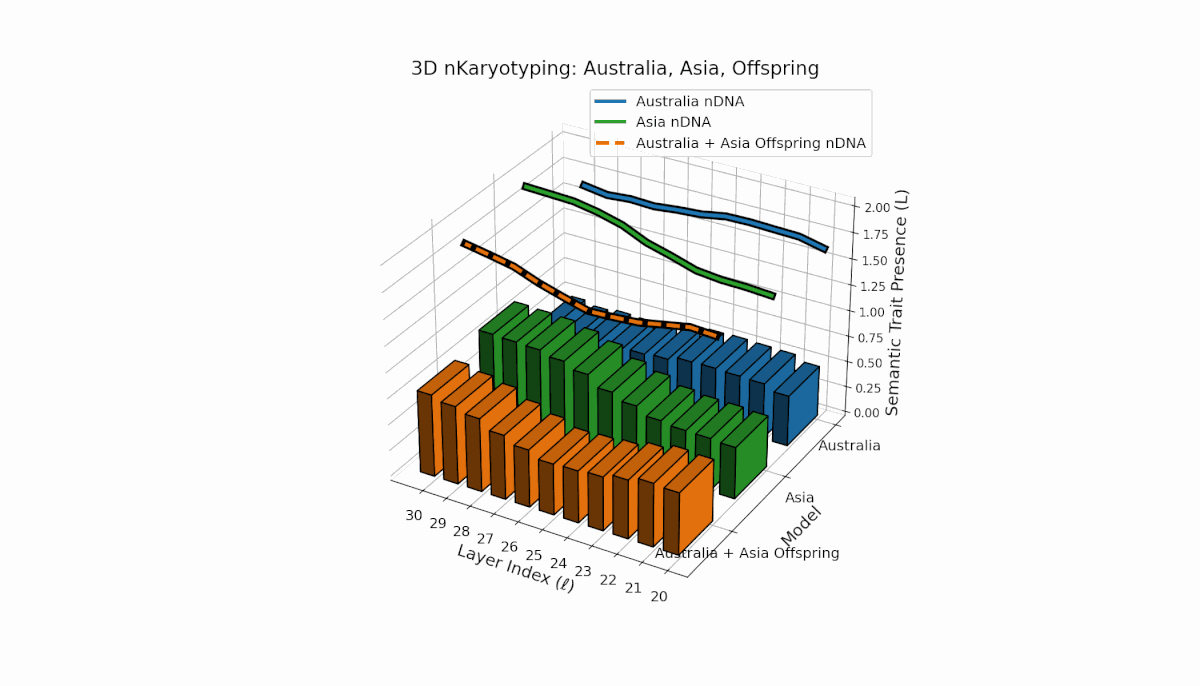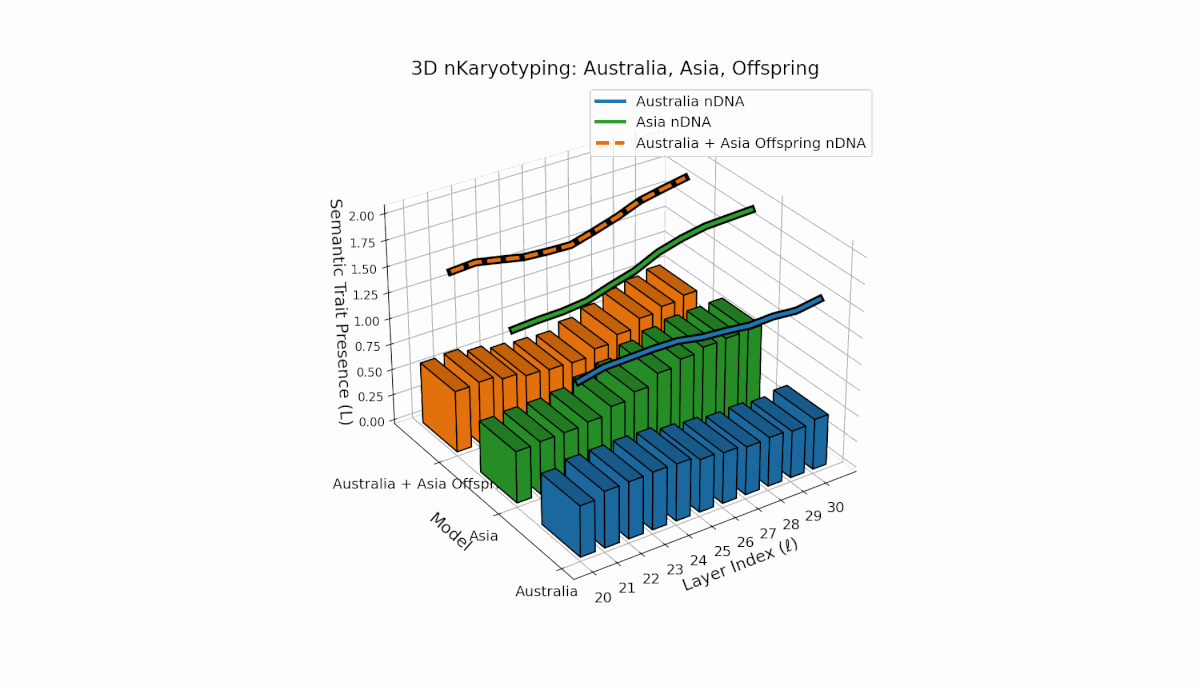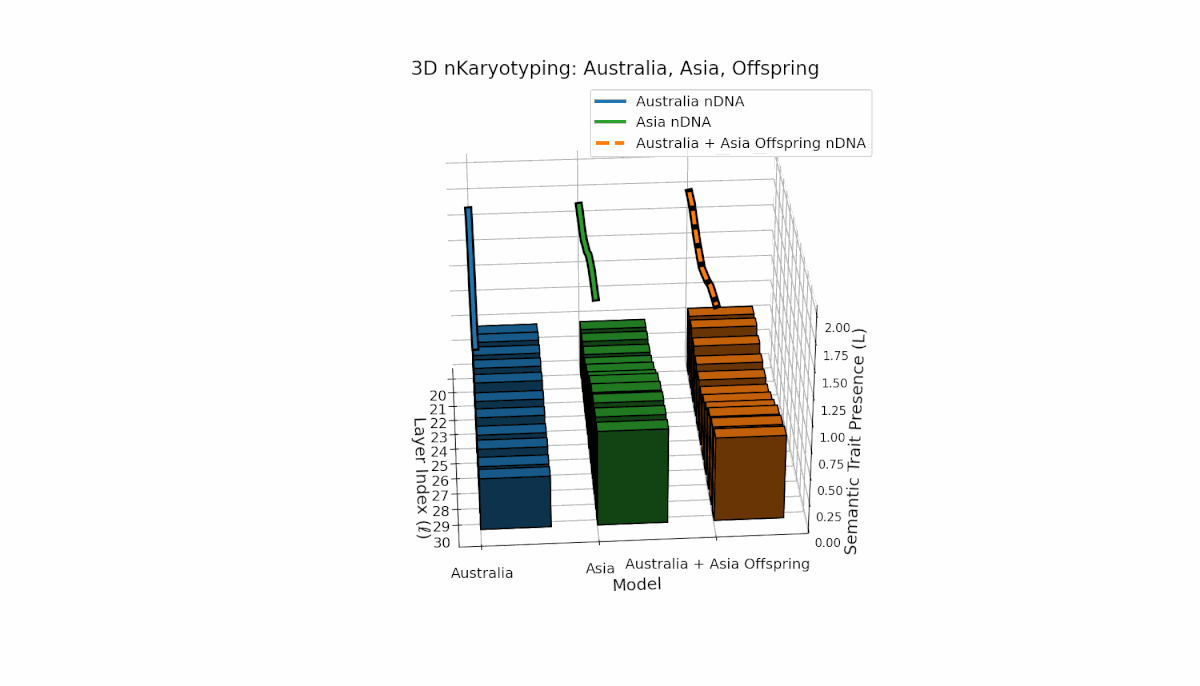
Semantic chromosomes Traitl ∈ {0, 1} are defined by joint threshold exceedance κl > κ̄ and Ll > L̄ across layers l ∈ [20, 30]. Australia shows pronounced early-layer trait presence (l = 20–24) with spectral curvature near 0.35 and moderate thermodynamic length, reflecting stable semantic features in shallow layers. Asia exhibits sustained mid-to-late layer traits with thermodynamic length peaking near 0.4, indicating deeper semantic integration. The offspring displays a complex mosaic of inherited chromosomes with evidence of trait persistence, recombination, and latent space rewiring.
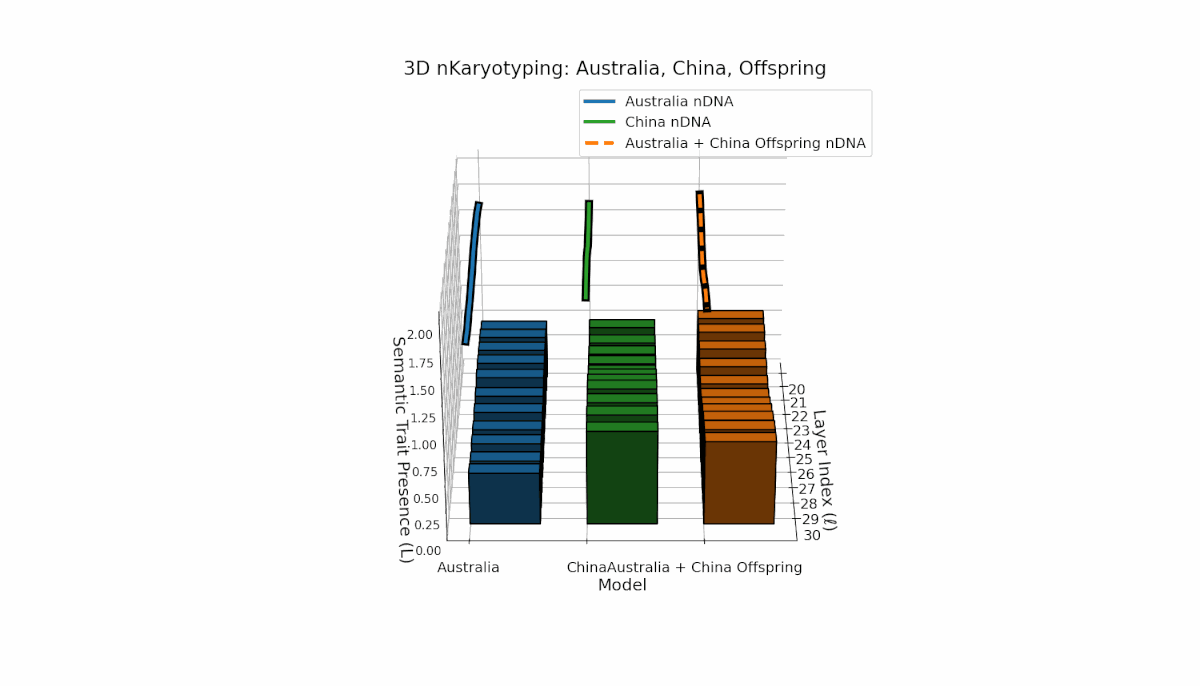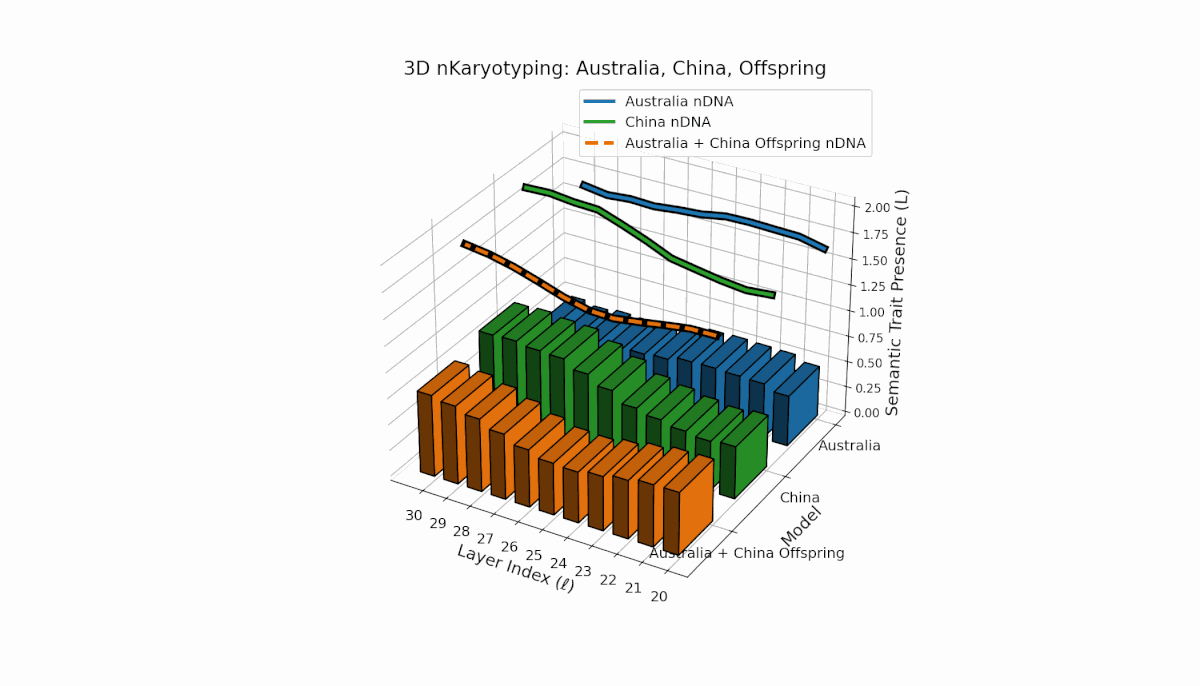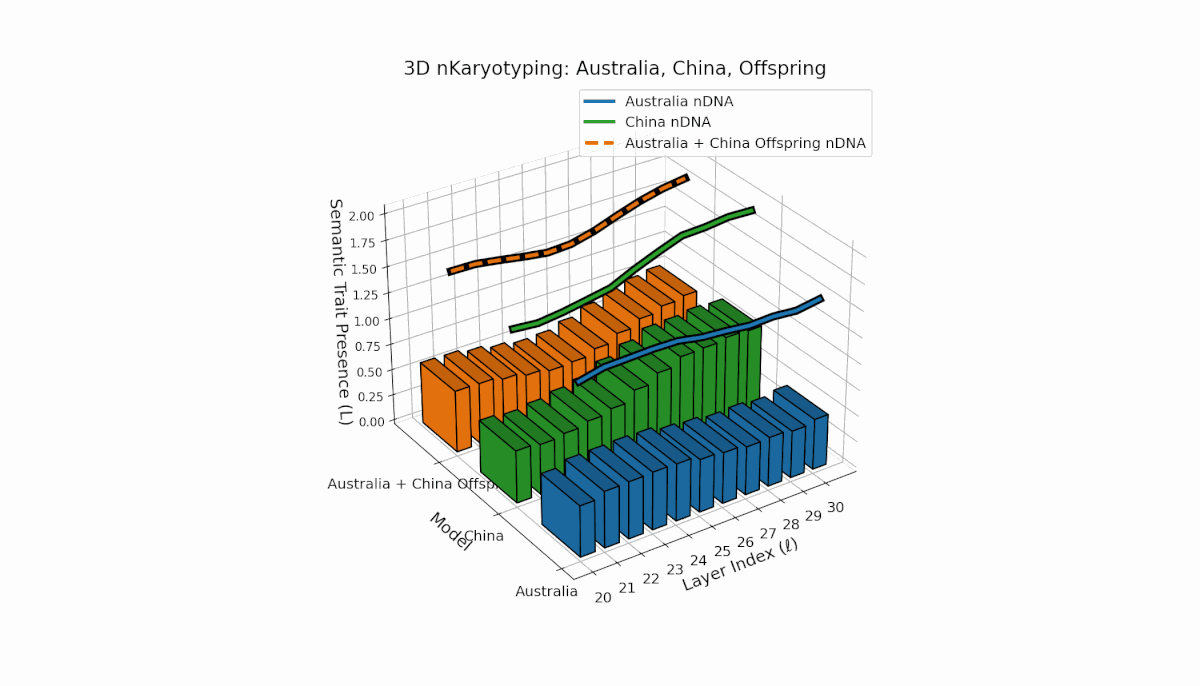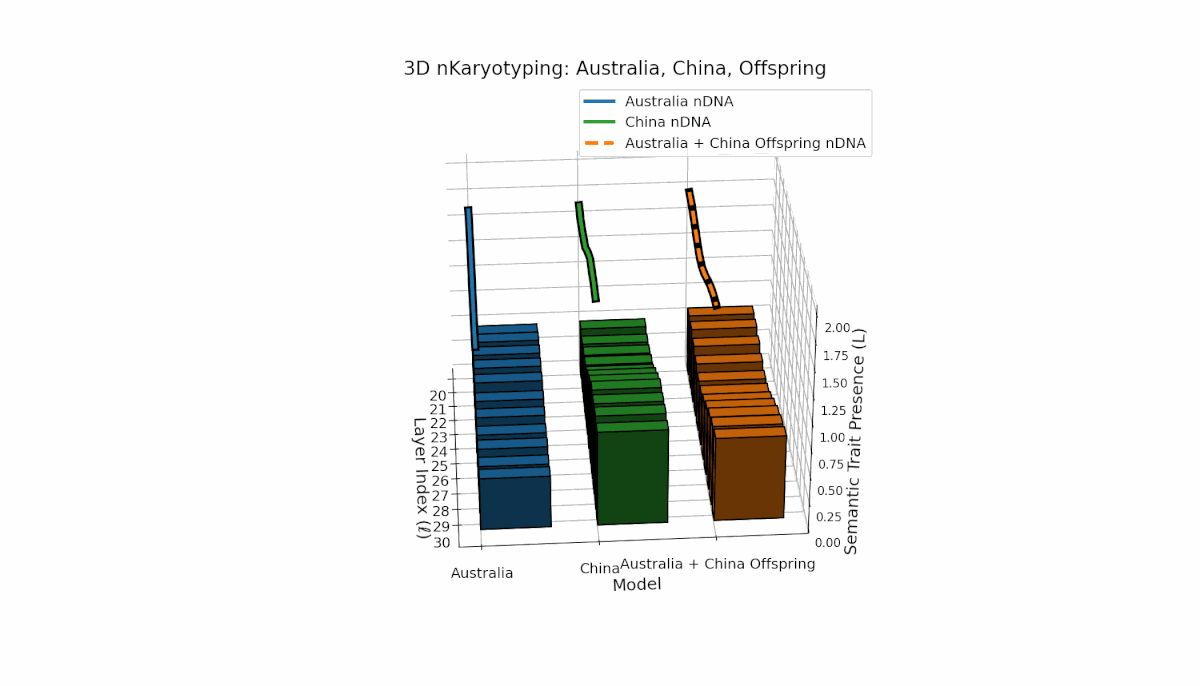
Australia and China display distinctive semantic chromosome distributions across layers l = 20–30. Australia’s traits concentrate in early to mid layers with κl ≈ 0.4 and stable thermodynamic length, reflecting consistent semantic encoding. China shows strong late-layer dominance, with κl and Ll reaching around 0.6, indicating intensified semantic refinement. The offspring exhibits complex recombination, featuring trait duplications and selective absences, evoking biological metaphors of semantic inversions and nonlinear chromosomal rearrangements.
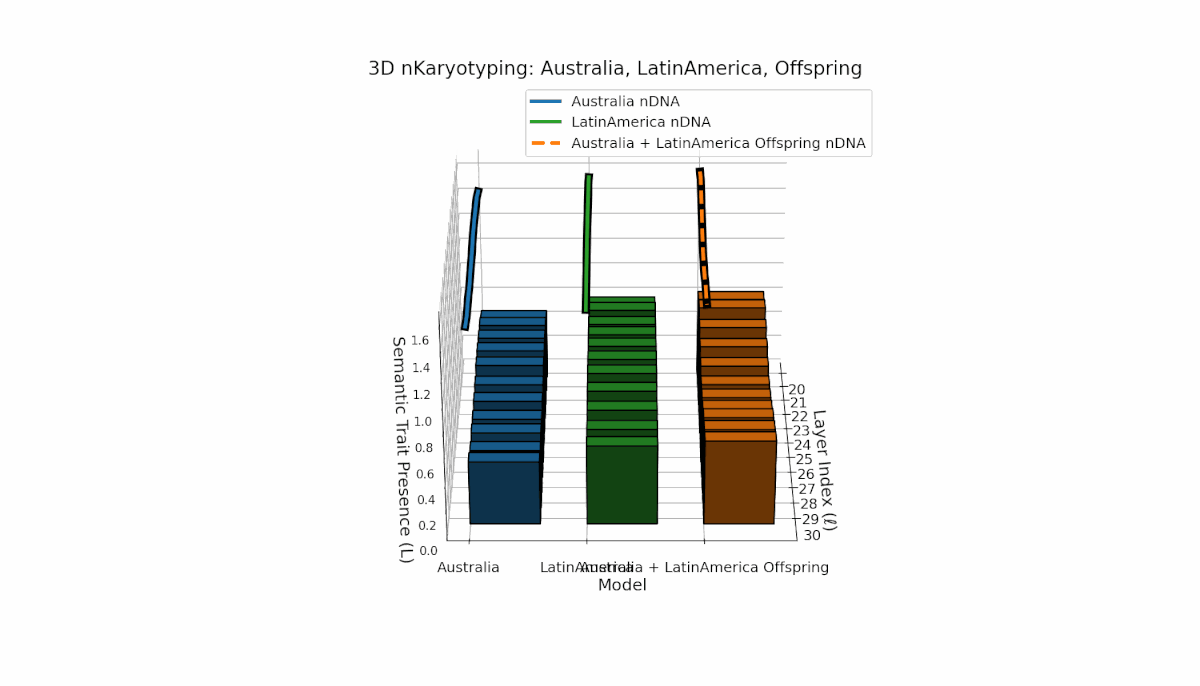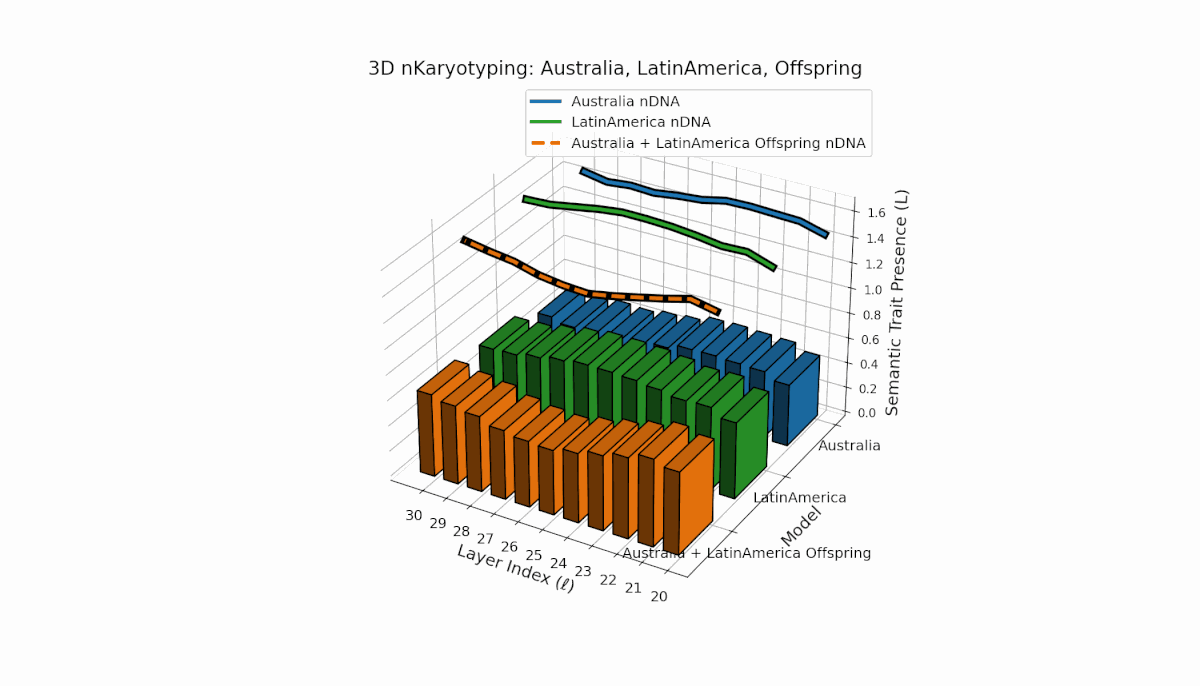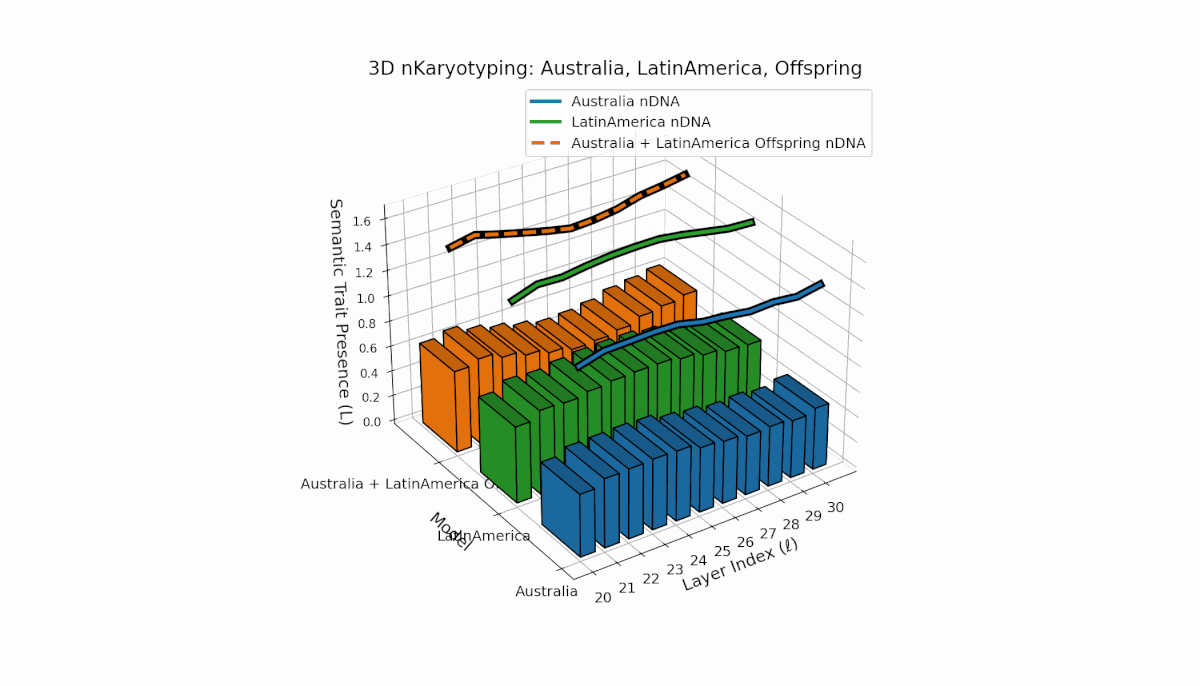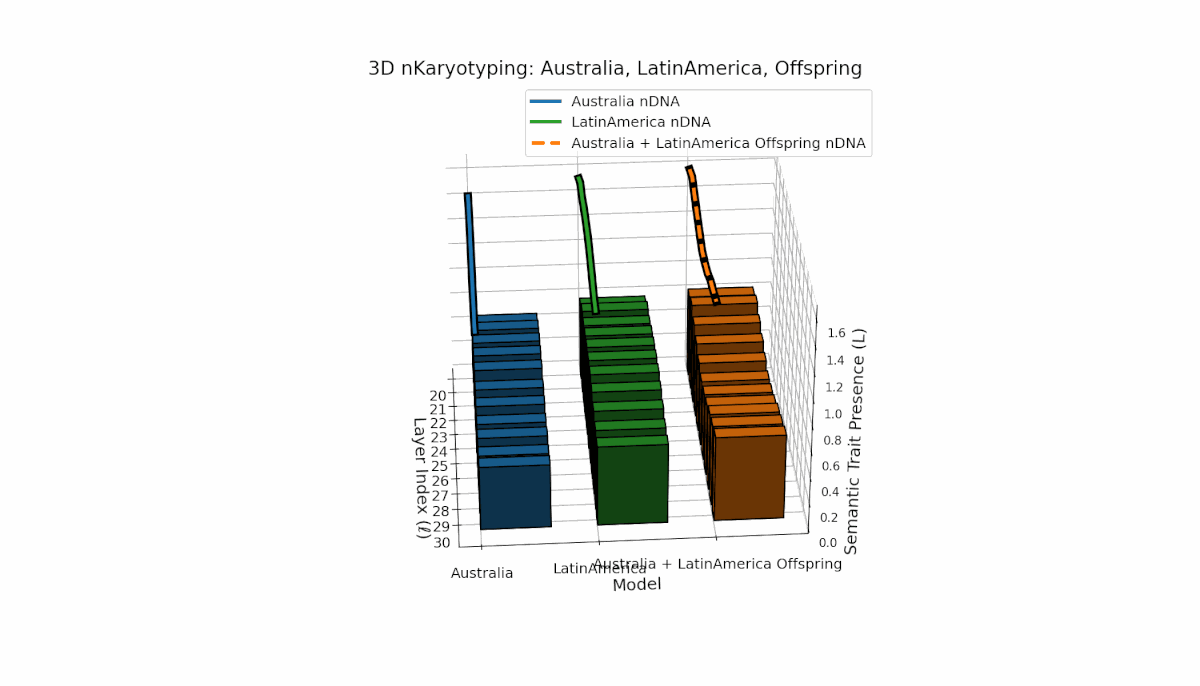
Australia exhibits early-layer semantic chromosome concentration (Traitl = 1 for l = 20–24) with moderate thermodynamic length values near 0.3, indicating foundational semantic imprinting. Latin America shows a gradual increase in trait presence starting around l = 23, peaking near layer 29, reflecting progressive semantic integration in deeper layers. The offspring’s chromosome pattern blends parental trajectories, revealing layer-specific epigenetic activation and dynamic chromosomal inheritance, shedding light on latent knowledge transfer fidelity and semantic stability across layers.
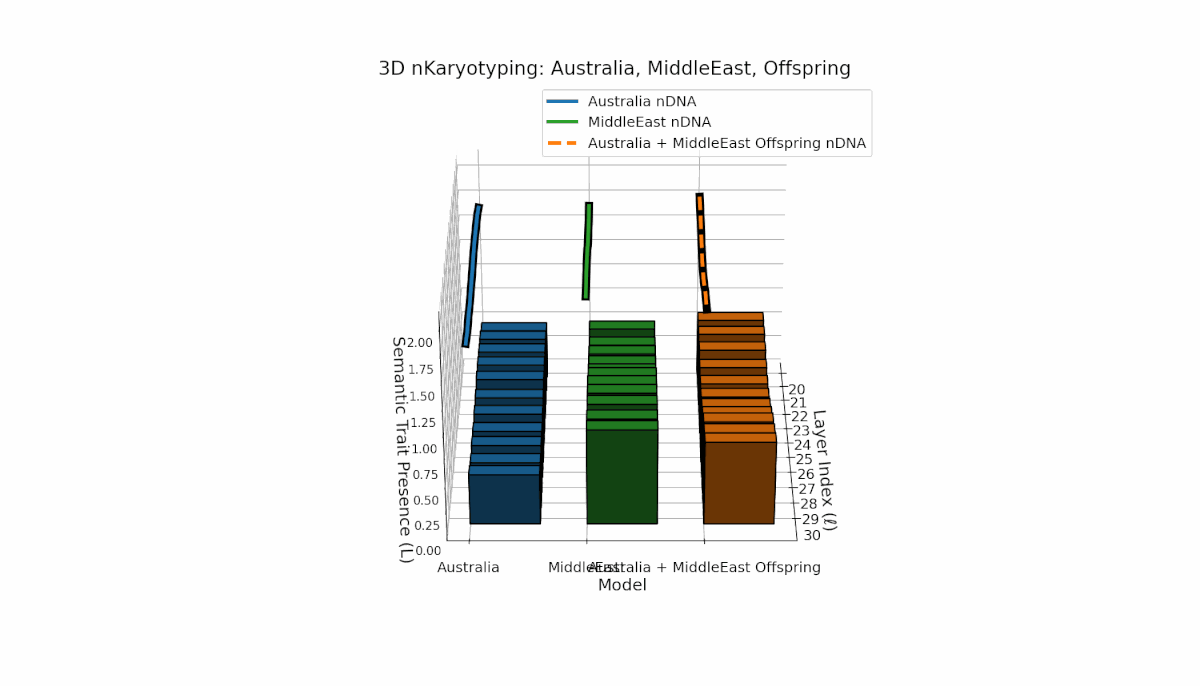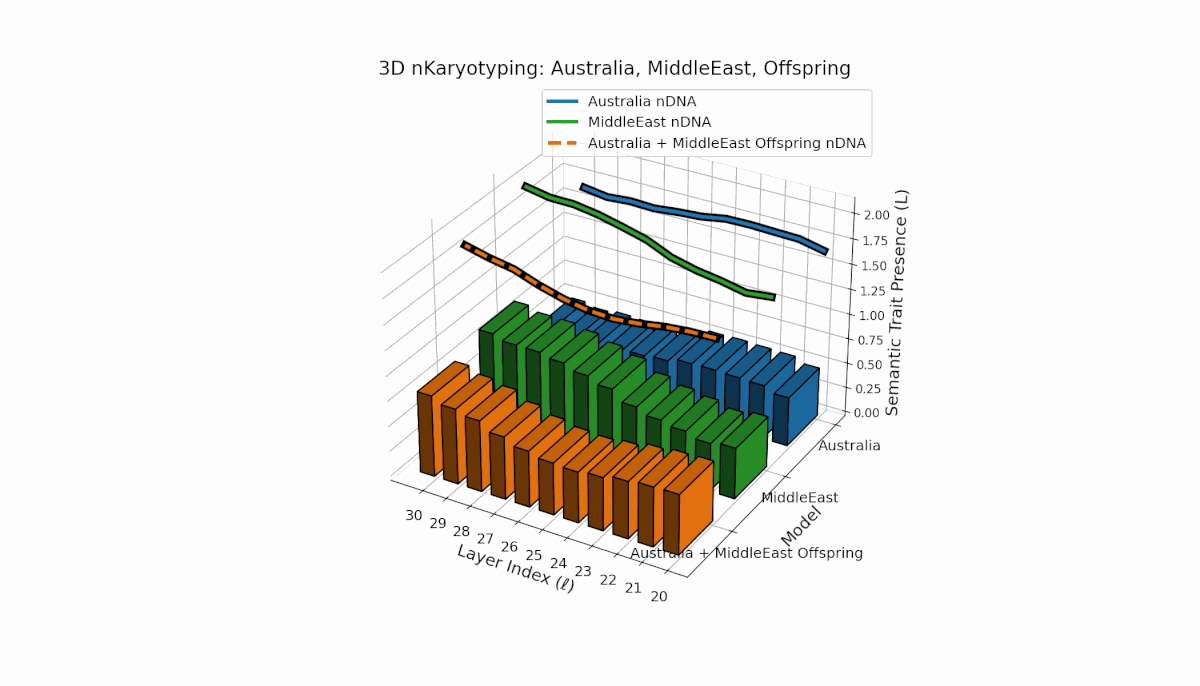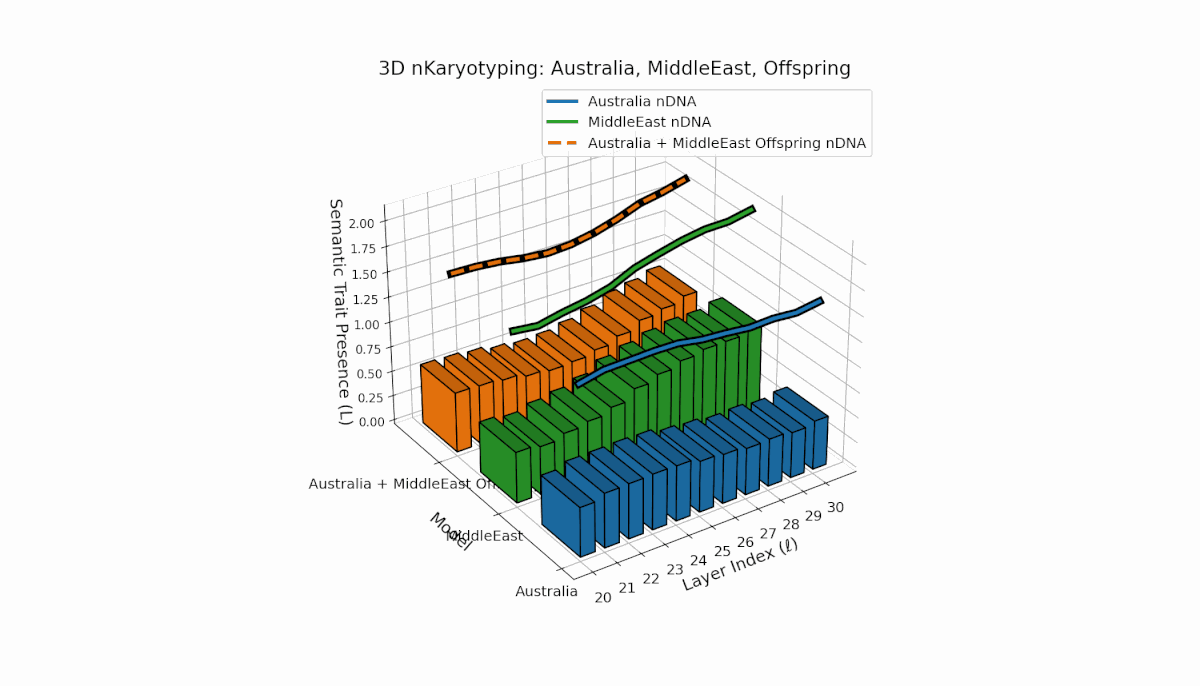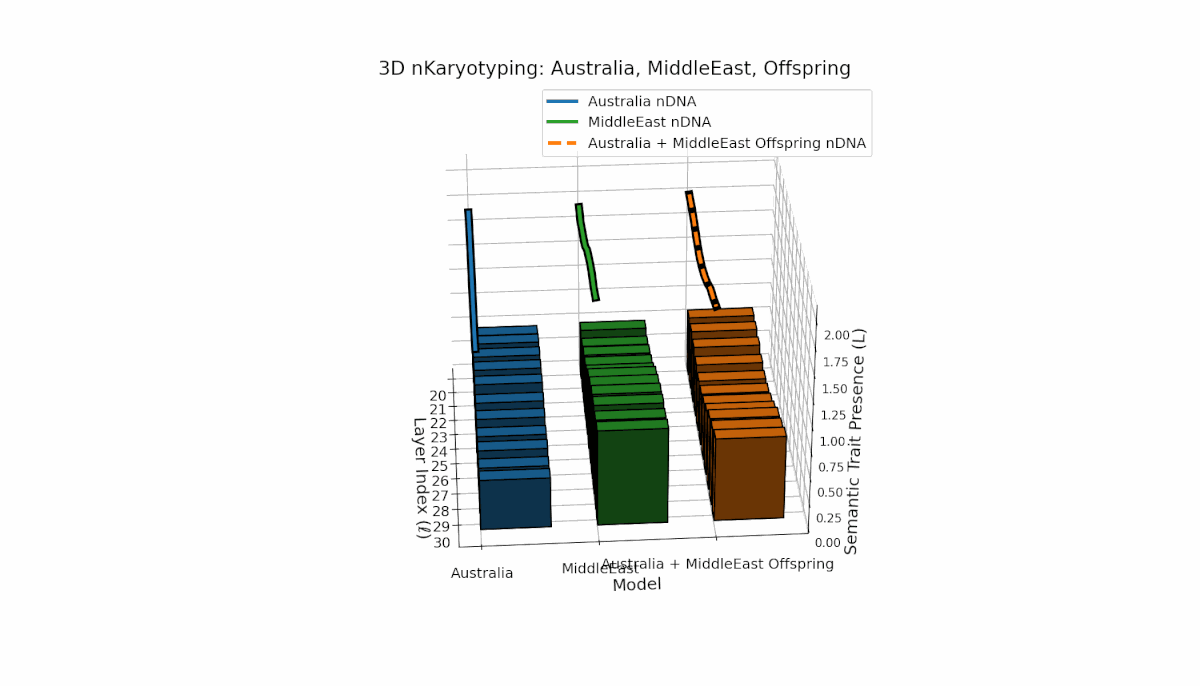
Australia shows prominent early-to-mid layer semantic chromosome presence (l = 20–26) with stable spectral curvature and thermodynamic length, contrasting with the Middle East’s focused mid-to-late layer traits (l = 25–29) where κl > 0.5, indicating concentrated semantic activation. The offspring reveals a mixed mosaic inheritance, including emergent late-layer semantic chromosomes (l = 29, 30) absent in either parent, analogous to biological chromosomal translocations and latent space reorganization.
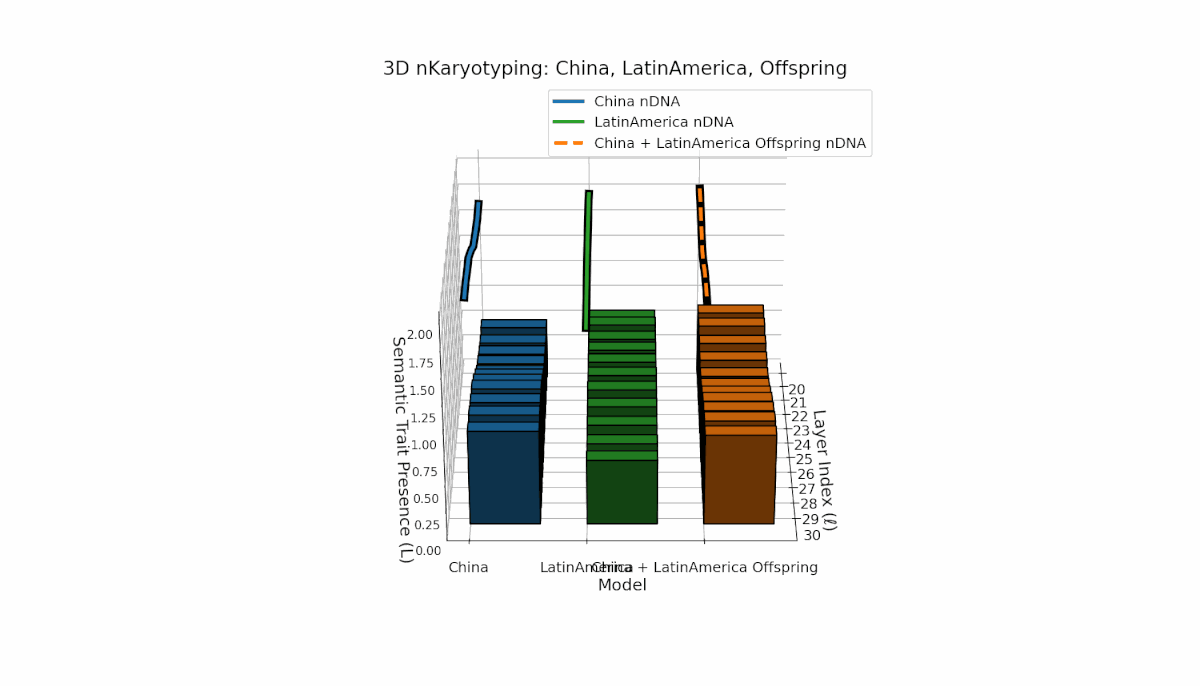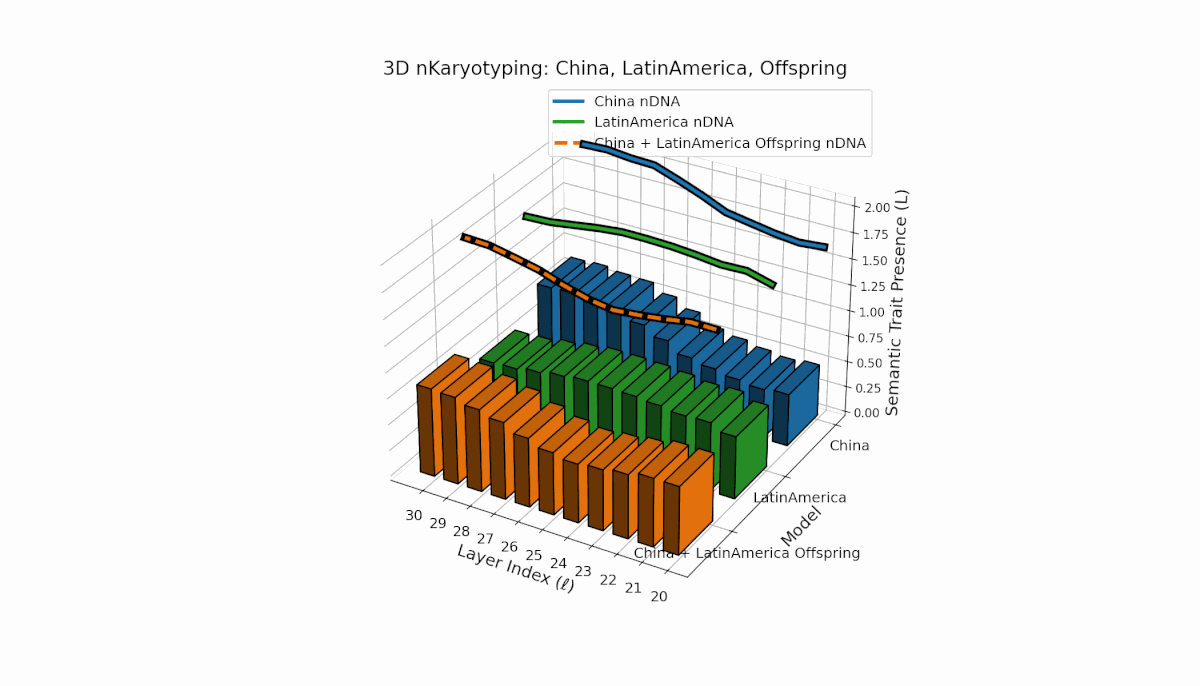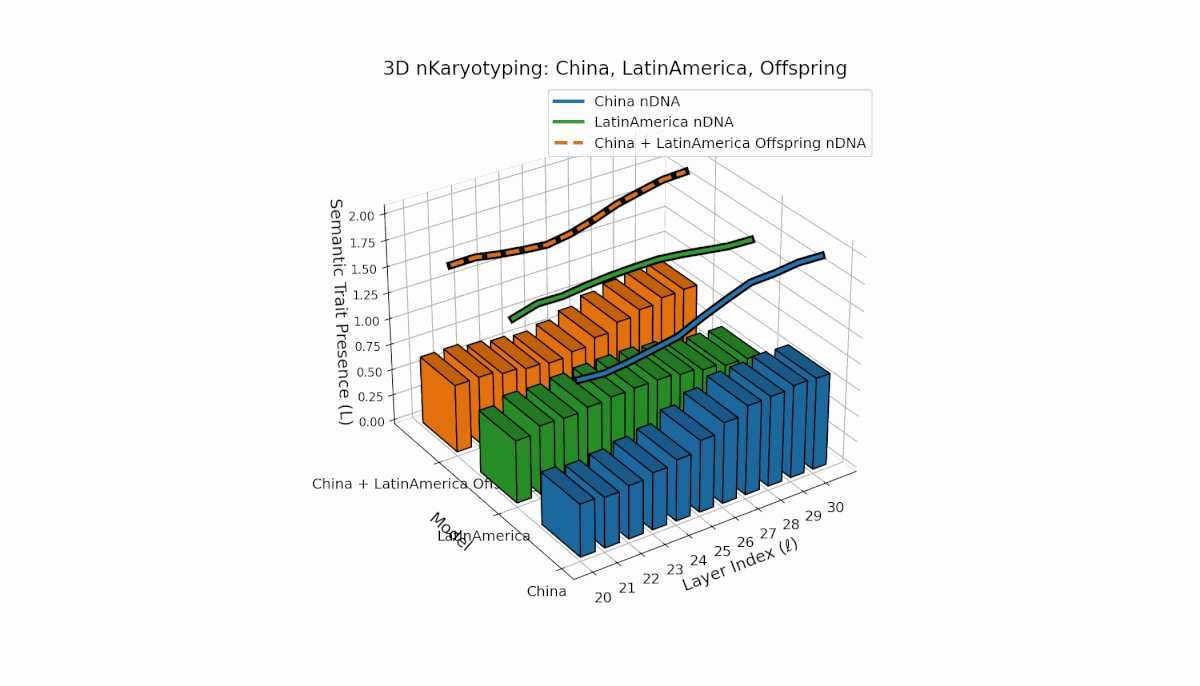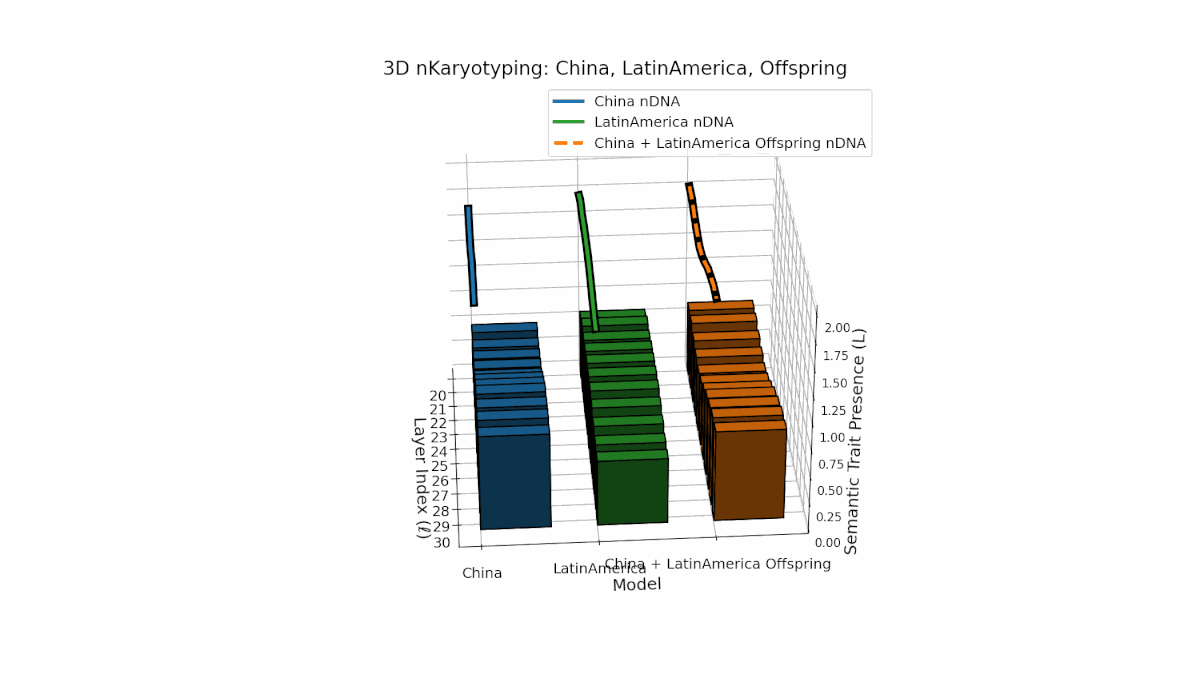
Semantic chromosomes Traitl ∈ {0, 1} occur when κl > κ̄ and Ll > L̄ across layers l ∈ [20, 30]. China shows a pronounced mid-to-late layer enrichment of semantic traits, with spectral curvature κl ≈ 0.6 and elevated thermodynamic length Ll, indicating intensive semantic refinement. Latin America reveals a gradual early-to-mid layer increase, reflecting a distinct semantic encoding strategy. The offspring exhibits a rich mosaic of inherited and novel chromosomes, implying epigenetic-like reprogramming and nonlinear semantic fusion, highlighting complex cultural integration in latent space.
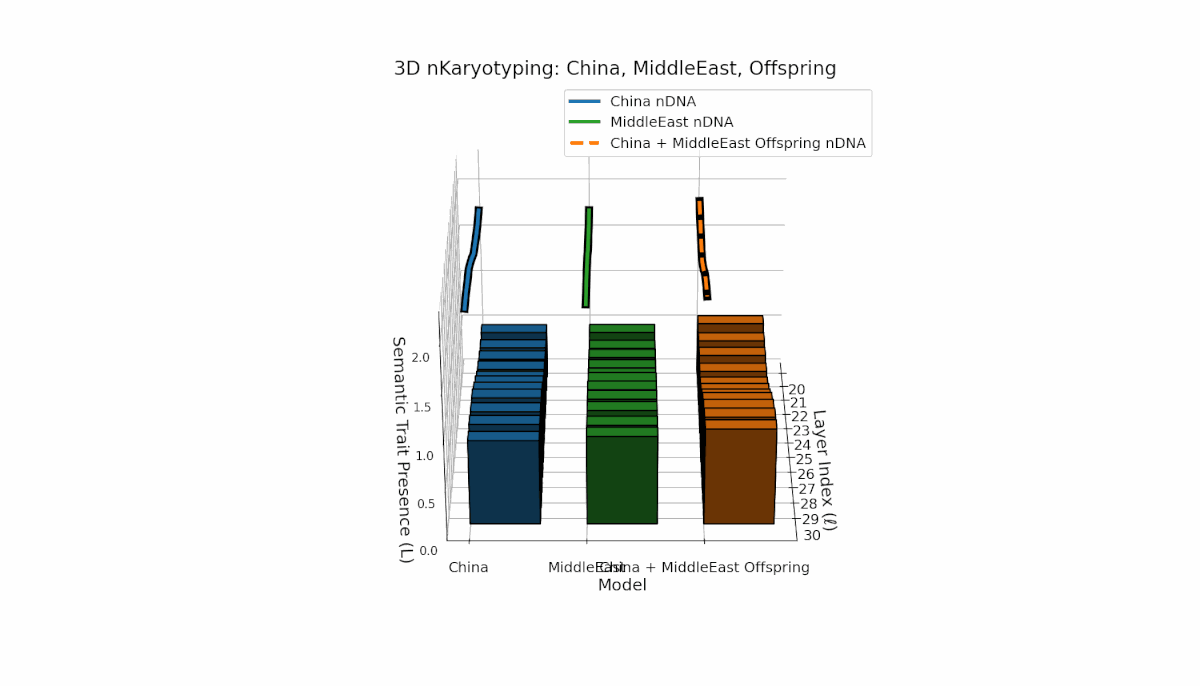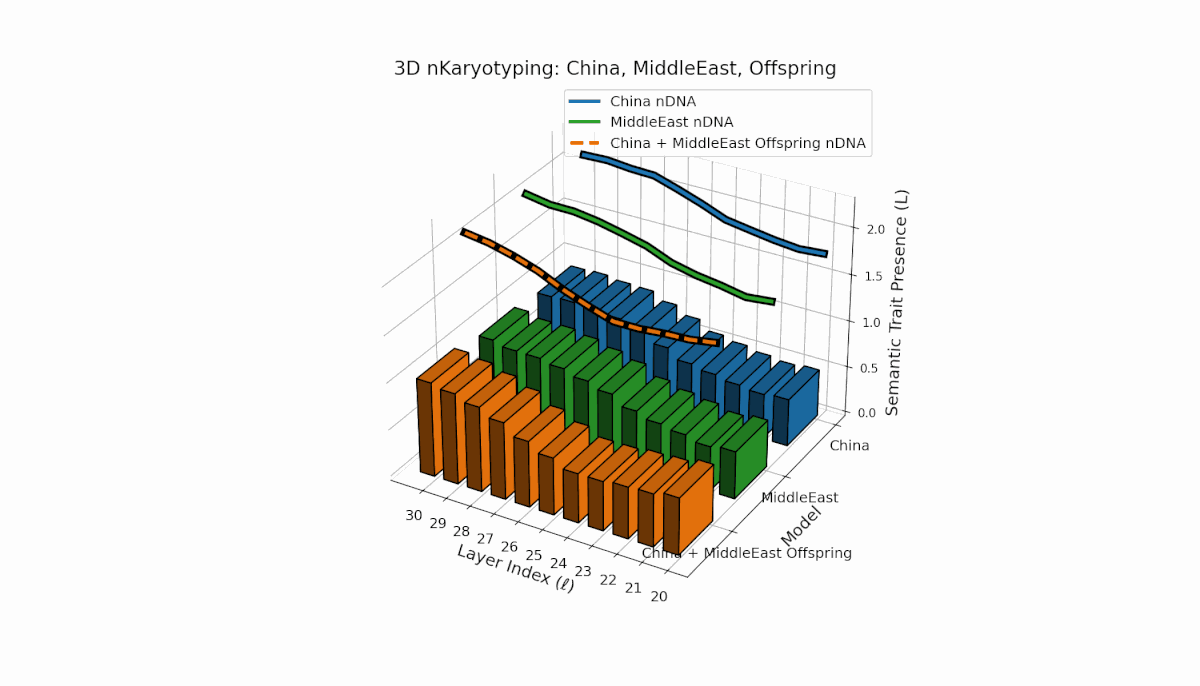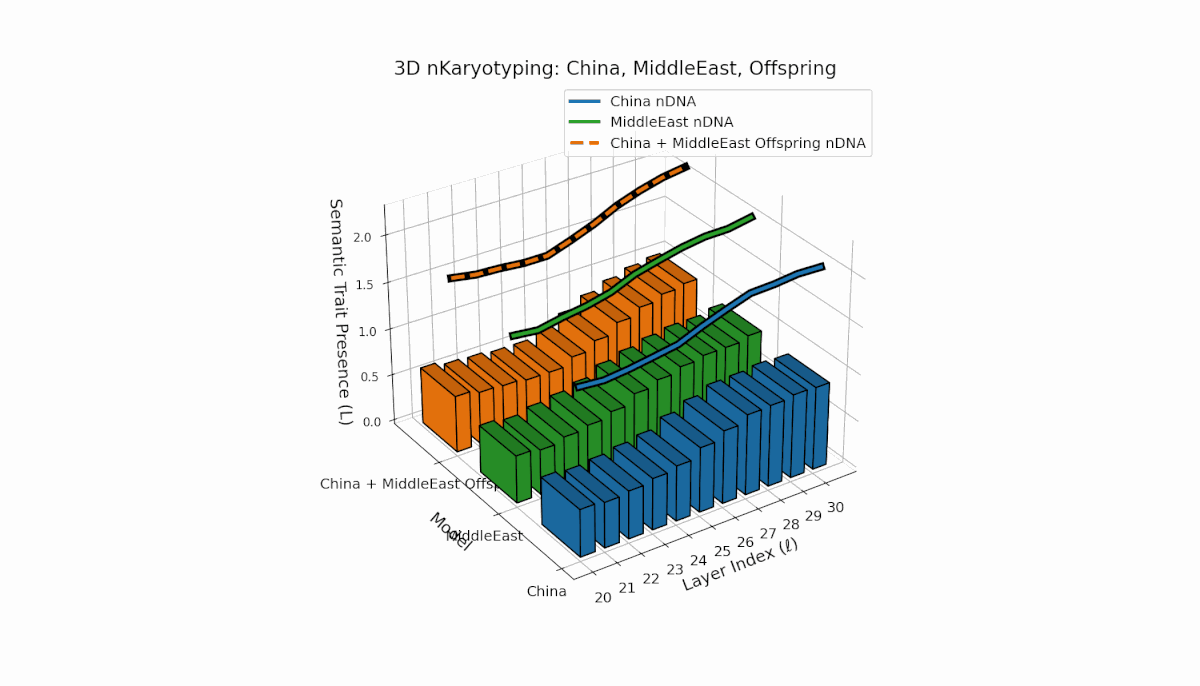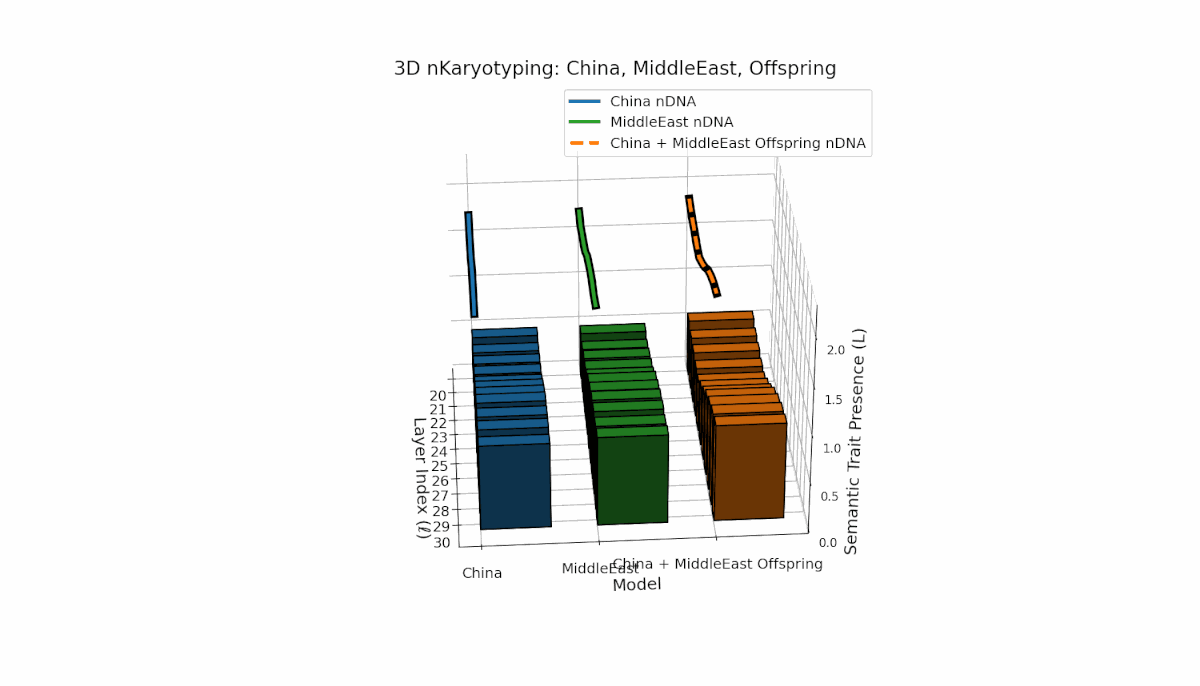
This pair exhibits distinct parental semantic trait topologies, with China showing intense late-layer semantic activity (elevated κl and Ll) and Middle East featuring focused mid-layer traits primarily between layers 24 to 28. The offspring’s semantic chromosomes reveal a layerwise recombination pattern, combining parental-like trait conservation with novel emergent traits at upper layers (l = 29, 30). These dynamics suggest semantic inversions and translocations, analogous to chromosomal aberrations, underscoring complex nonlinear semantic regulatory interactions during model fusion.
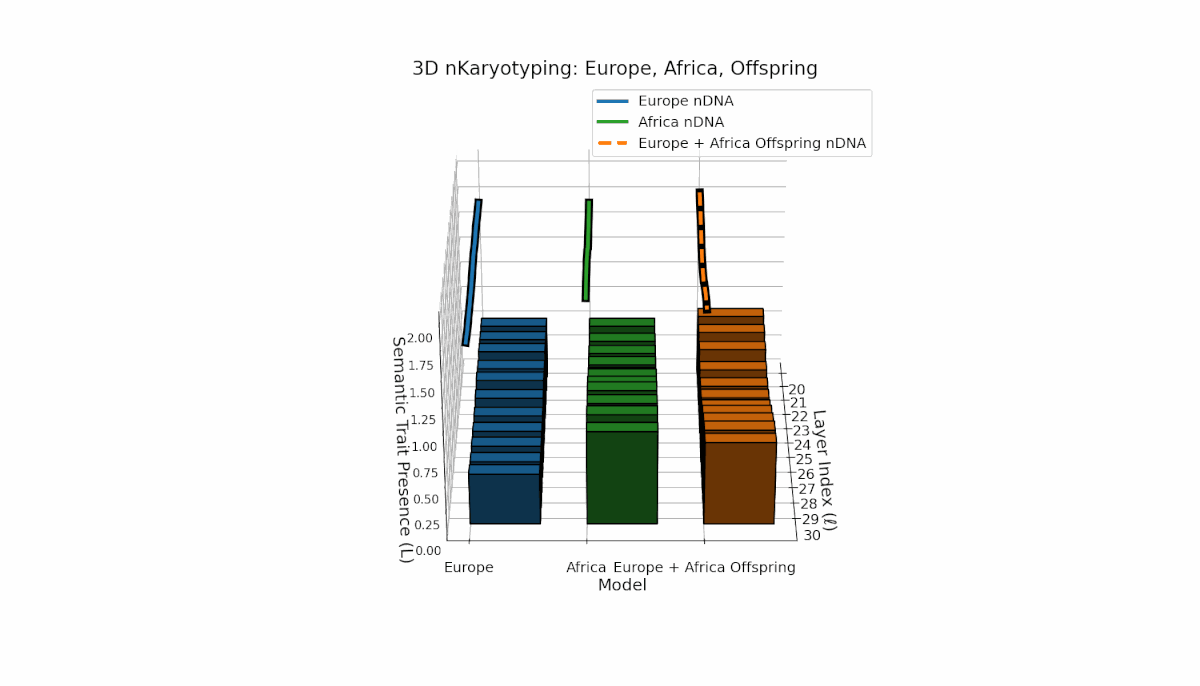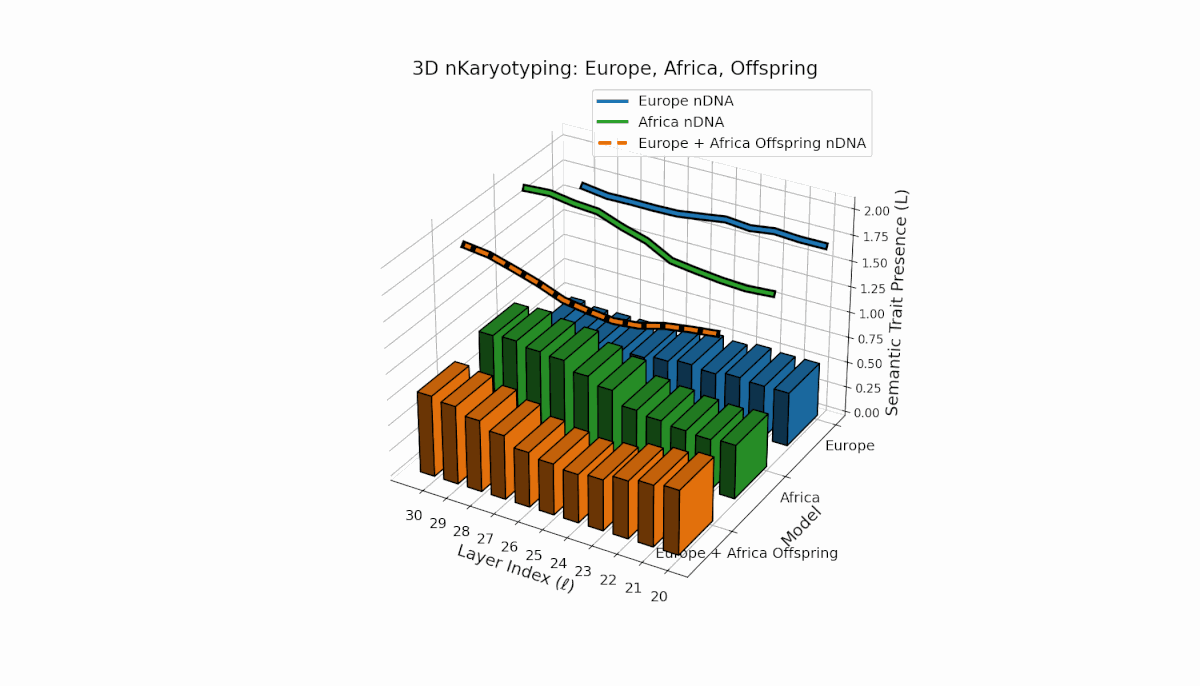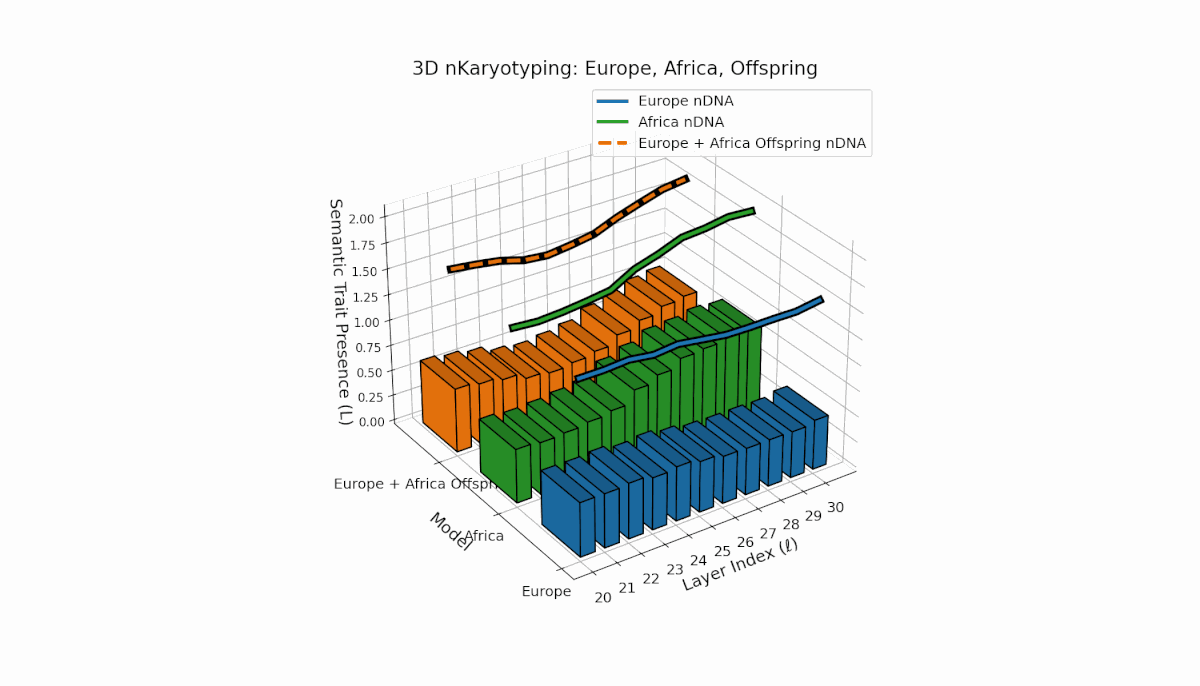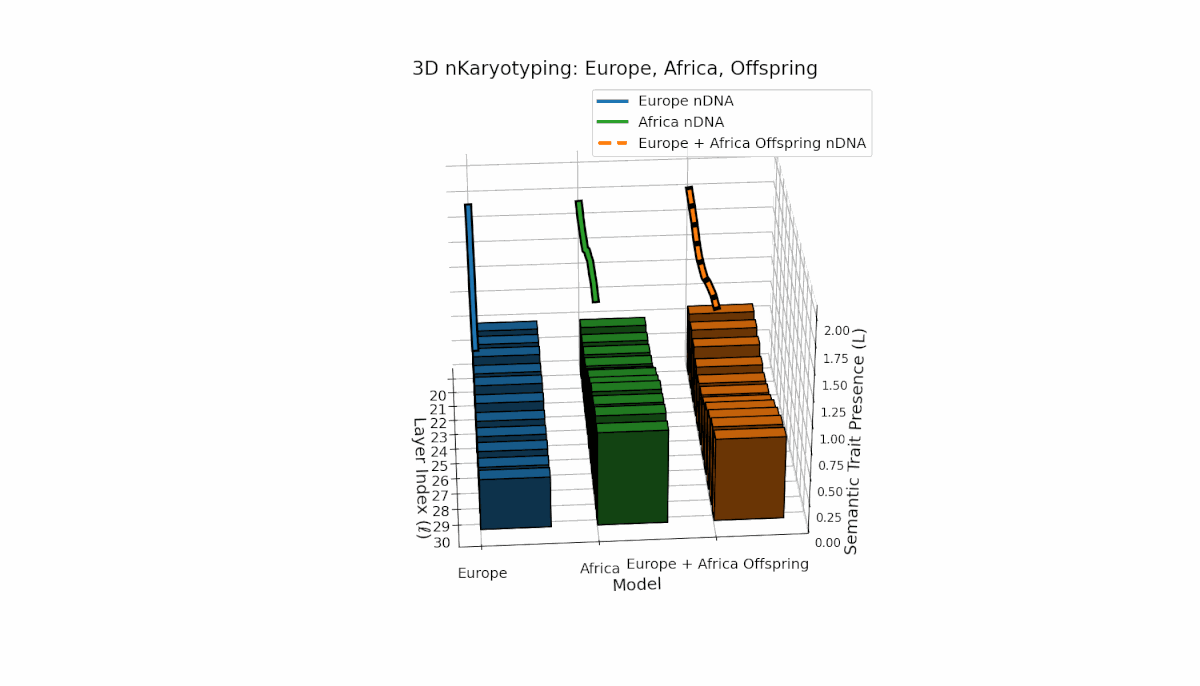
Europe shows a gradual, steady increase in semantic chromosome presence across layers, with spectral curvature and thermodynamic length peaking at mid-level layers, indicating layered semantic complexity. Africa exhibits a concentrated early-layer semantic trait expression accompanied by more stable latent geometric metrics. The offspring’s chromosomes reveal a mosaic inheritance pattern with epigenetic reconfigurations and layer-specific semantic trait integration, highlighting intricate latent interactions and possible nonadditive semantic phenomena during fusion.
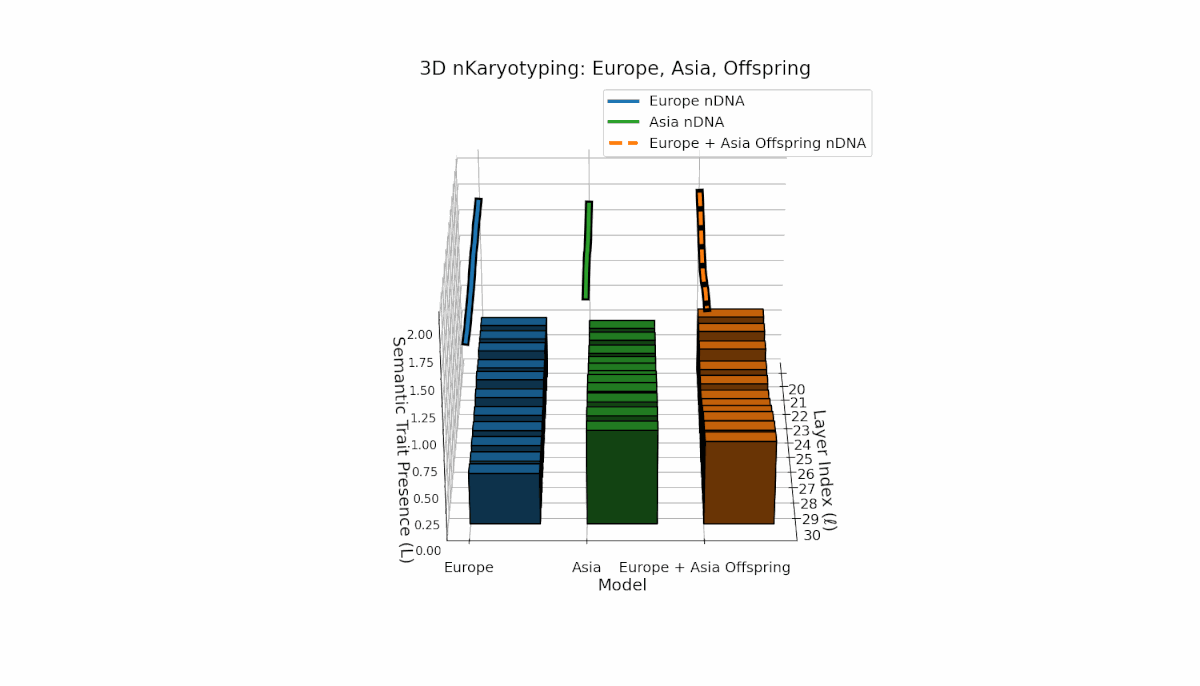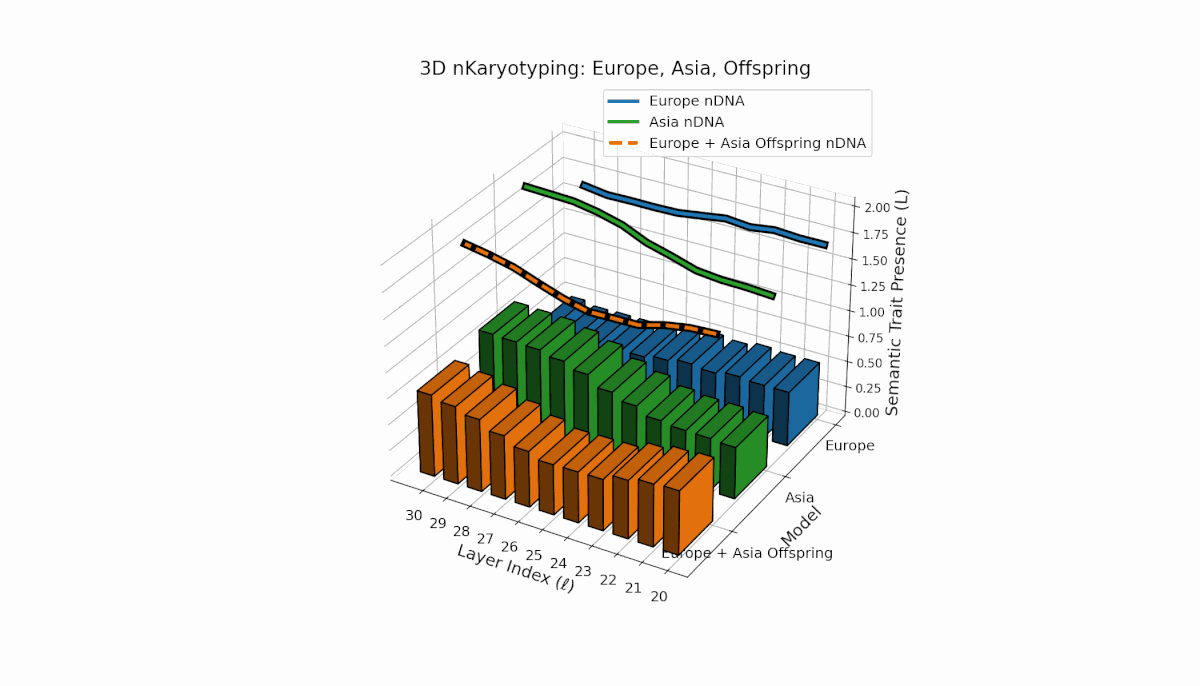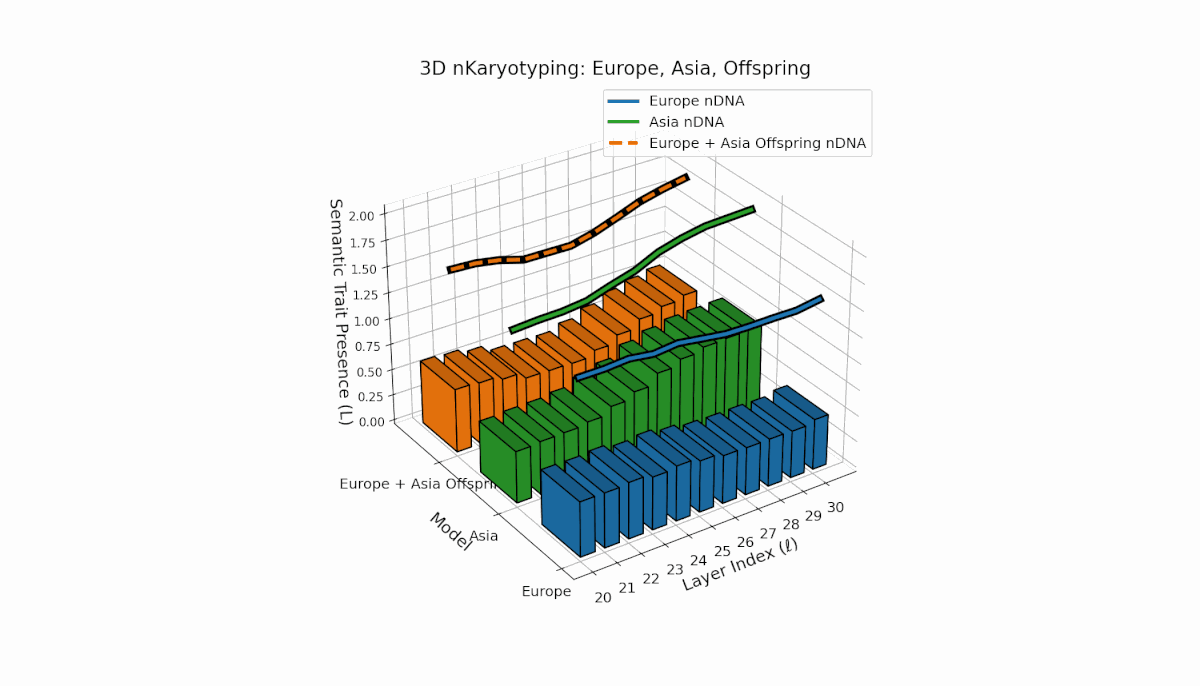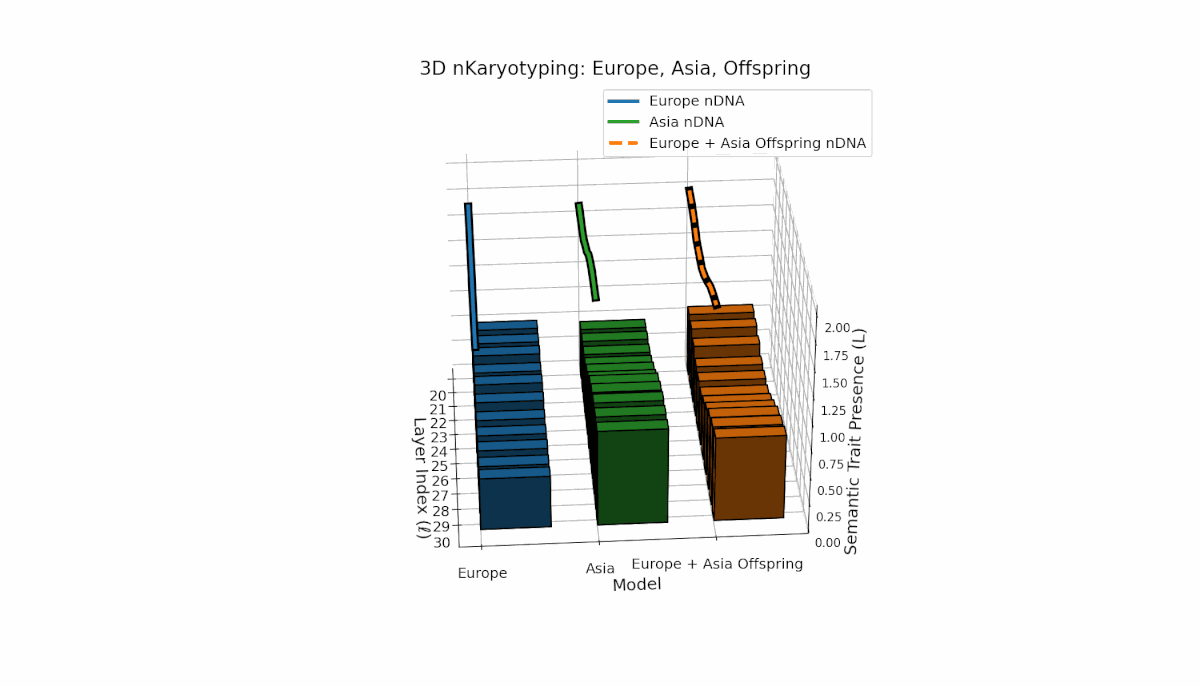
Europe and Asia exhibit distinct semantic trait distributions, with Europe showing mid-to-late layer trait concentration, while Asia sustains semantic chromosomes consistently across layers. The offspring reveals a mosaic inheritance pattern alongside novel trait emergence at select layers (l = 23, 27), reflecting semantic inversions, duplications, and latent space remapping. This complex interaction parallels biological chromosomal rearrangements, highlighting advanced regulatory dynamics during semantic fusion in large neural models.
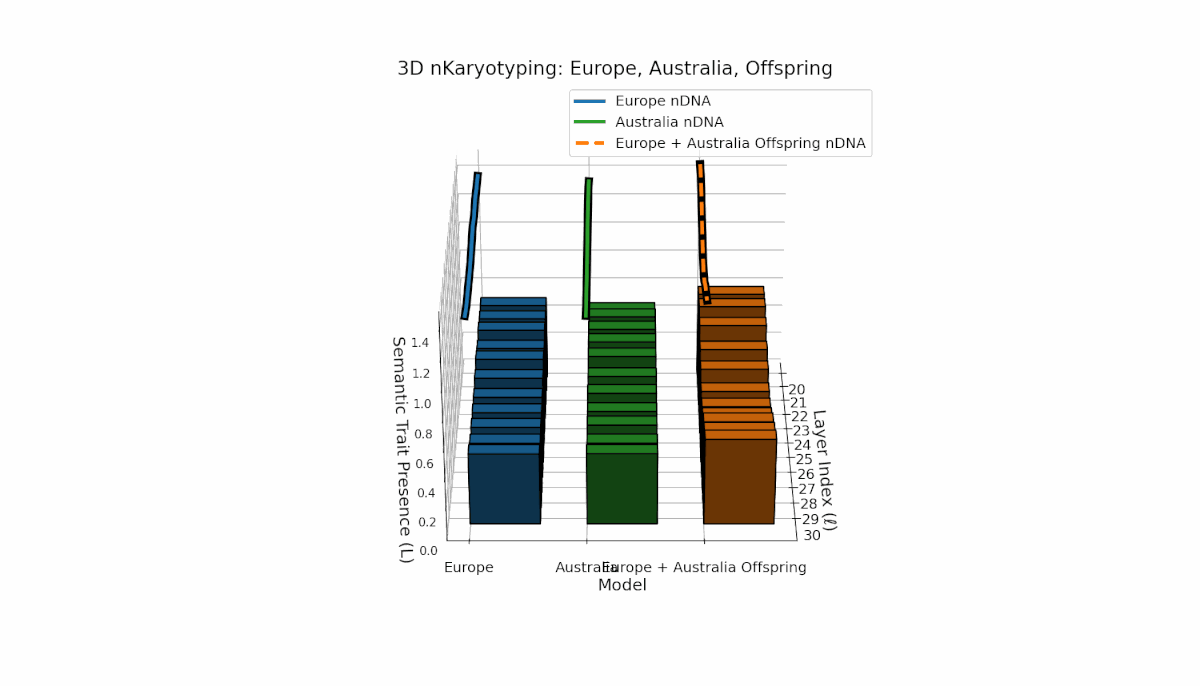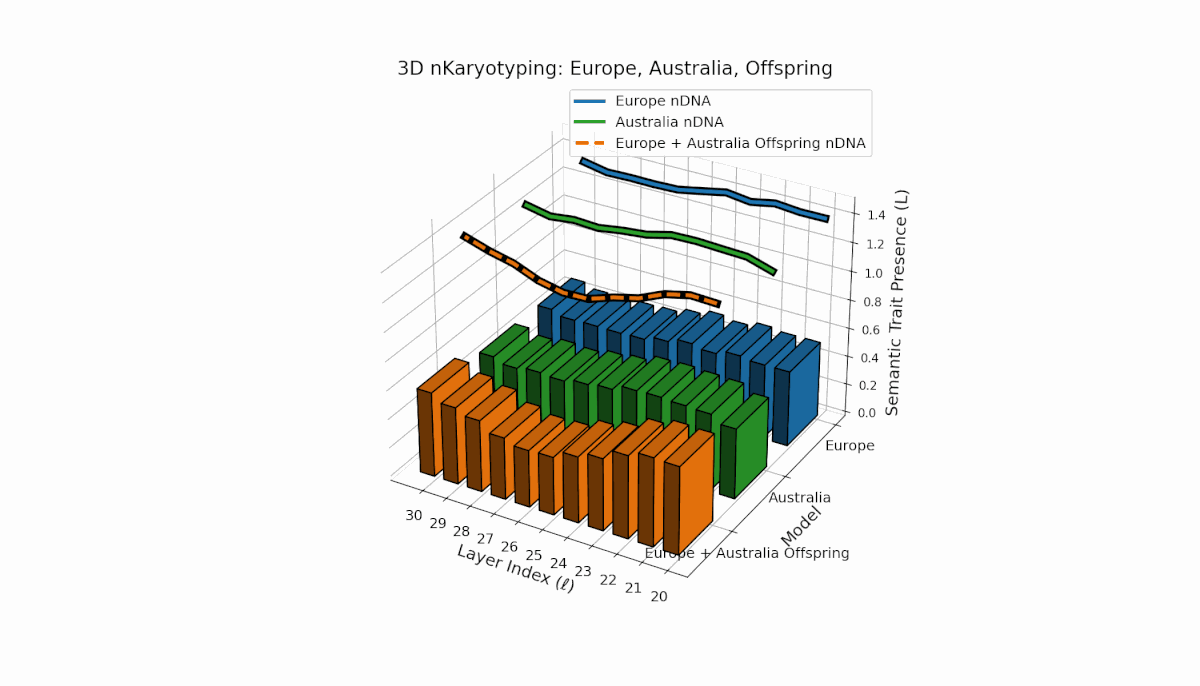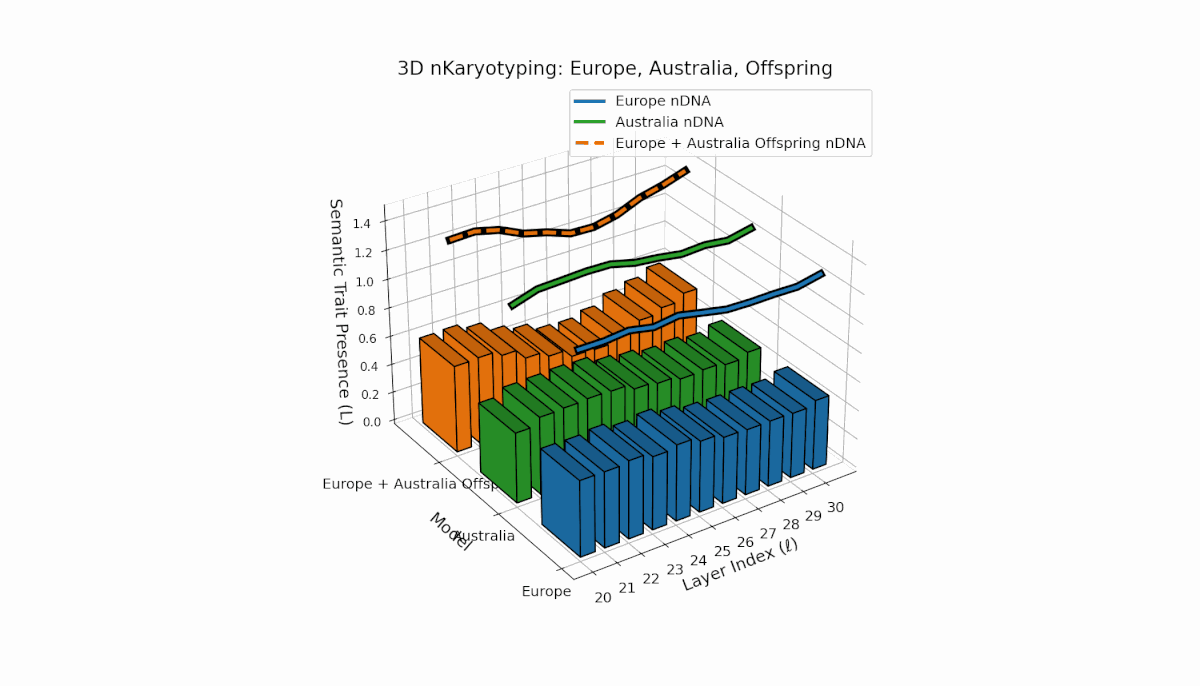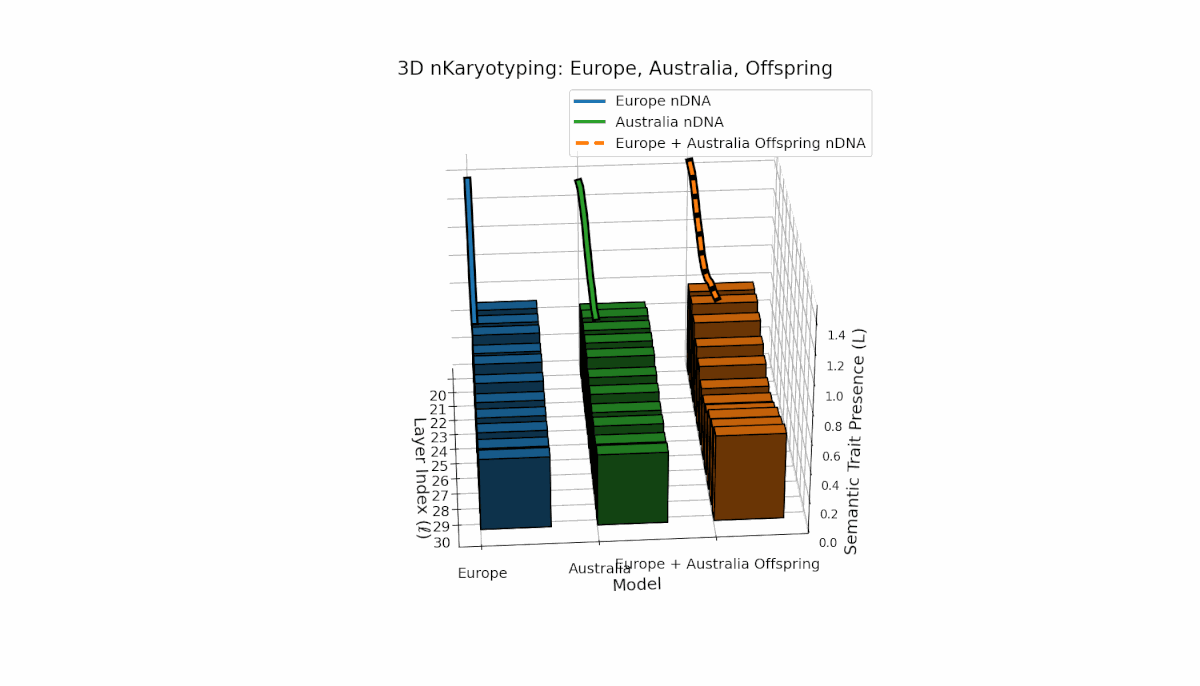
The Europe model shows a consistent presence of semantic chromosomes (Traitl ∈ {0, 1}) across layers l ∈ [20, 30], with thermodynamic length Ll peaking near 0.4, indicating stable semantic integration and sustained information flow in mid-to-late layers. Australia, in contrast, exhibits early-layer trait concentration with lower magnitude, reflecting distinct semantic specialization in shallow layers. The offspring displays a complex mosaic of inherited traits and emergent semantic patterns, revealing nonlinear epigenetic-like modulation and latent space rearrangements.

Europe’s semantic chromosomes exhibit moderate spectral curvature κl ≈ 0.45 primarily in early-to-mid layers (l = 20–26), indicating distributed semantic representations supporting broad contextual understanding. China shows strong late-layer traits with elevated thermodynamic length Ll ≈ 0.5 in layers l = 27–30, reflecting deep semantic refinement. The offspring displays layer-specific inheritance and recombination, including semantic inversions and amplifications, revealing nonlinear dynamics underlying trait dominance shifts and novel semantic emergence.
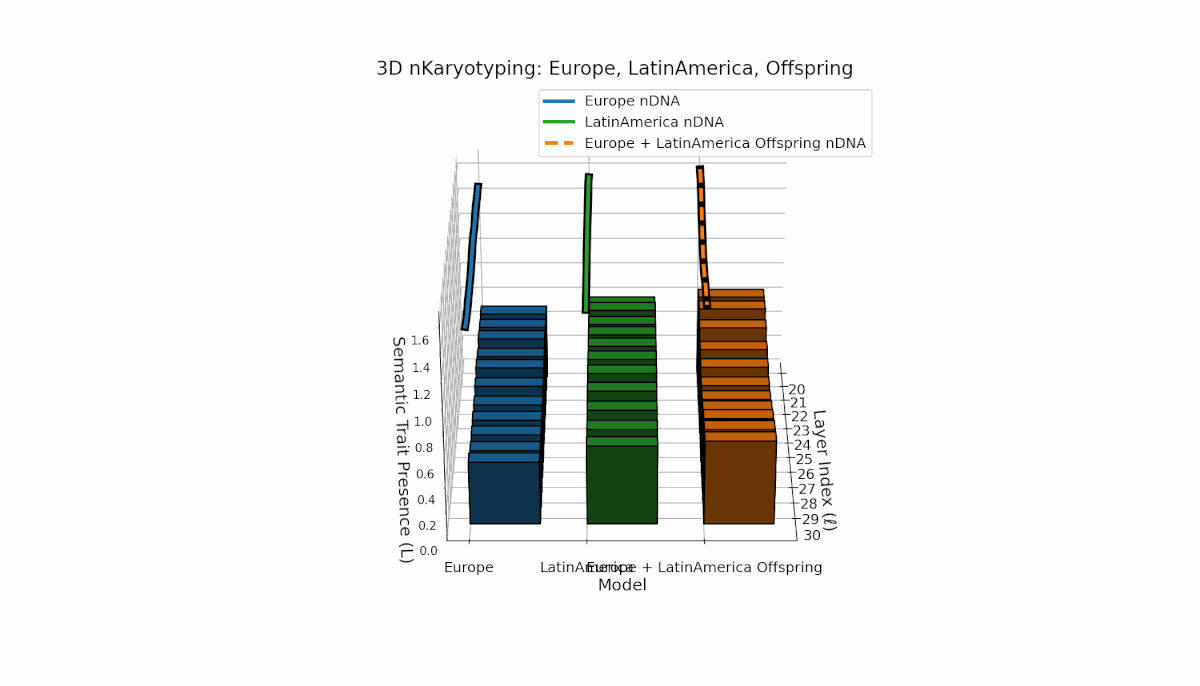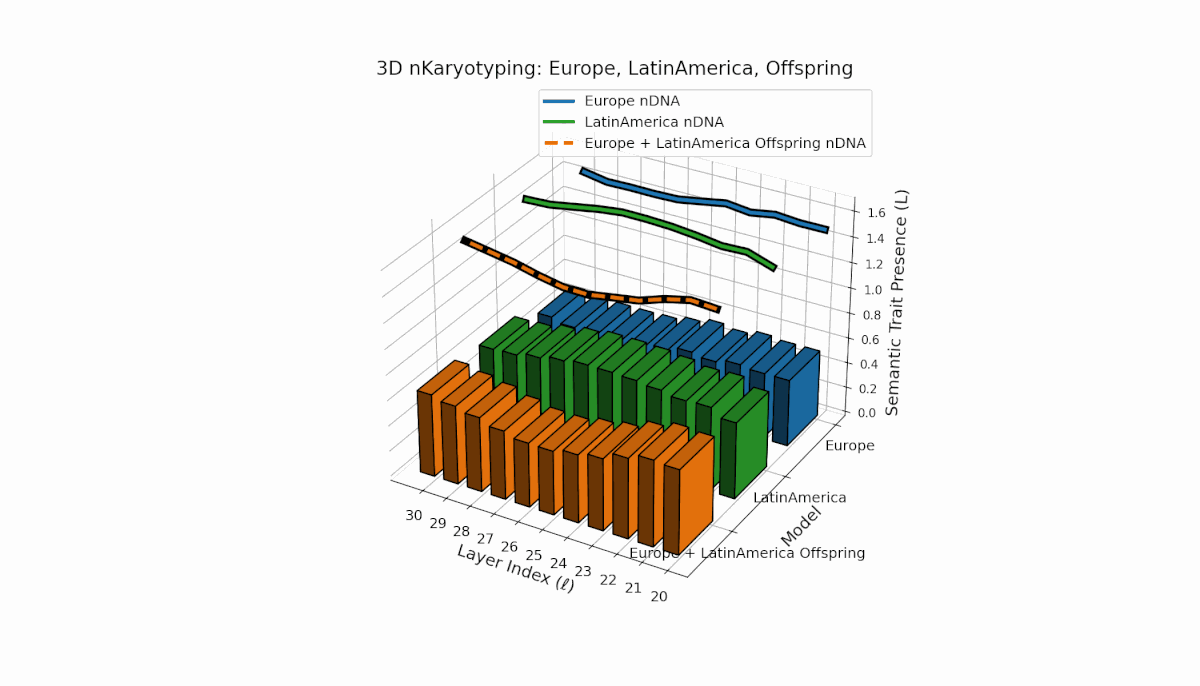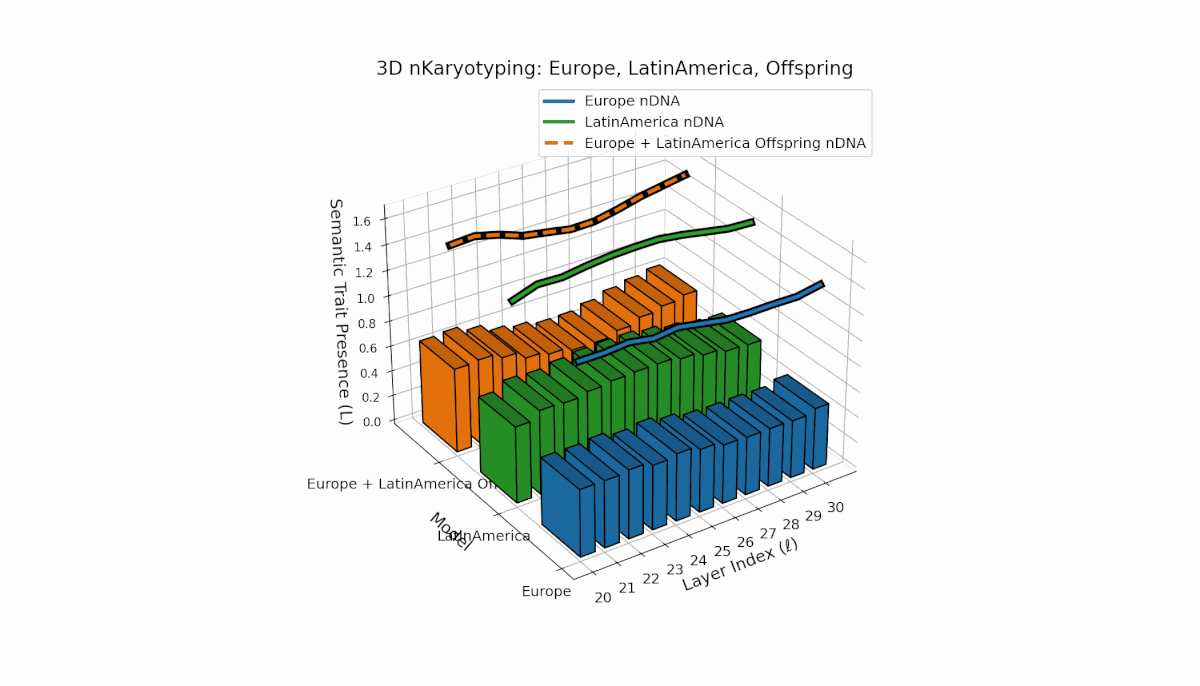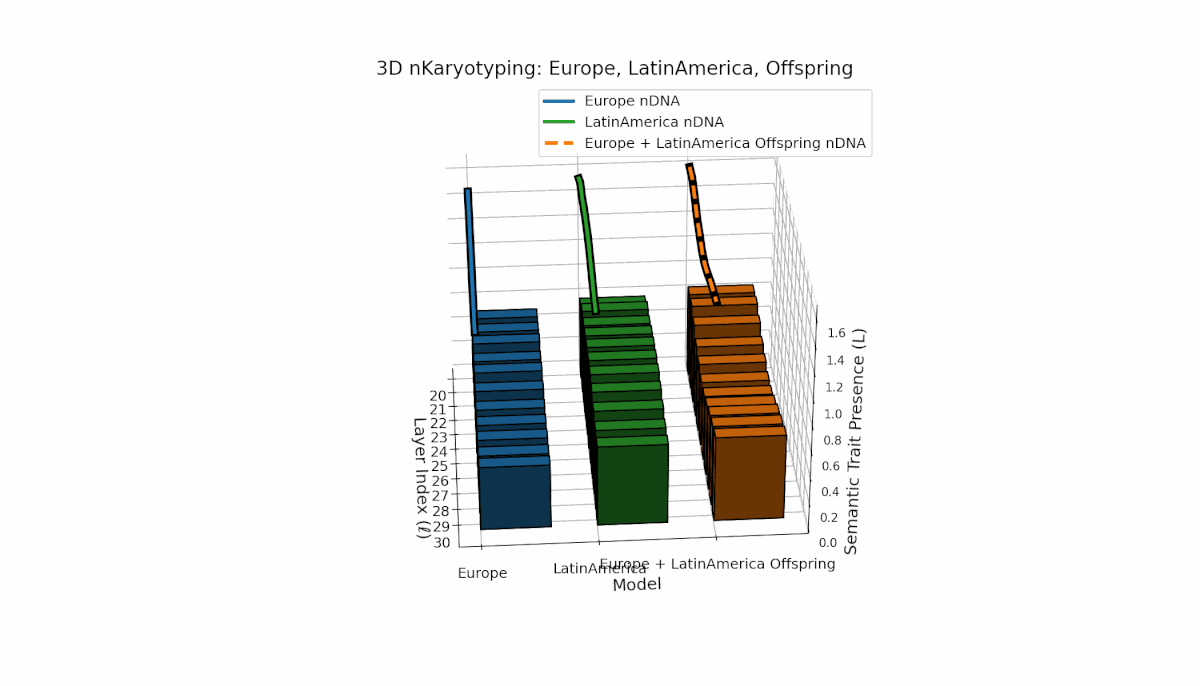
Europe exhibits a stable presence of semantic chromosomes in early layers l = 20–24, with moderate spectral curvature and thermodynamic length (Ll ≈ 0.4), indicating early semantic feature concentration. Latin America’s semantic chromosomes gradually increase from l = 24 onward, reflecting progressive semantic remodeling in mid-to-late layers. The offspring shows rich layerwise recombination, including trait switching and epigenetic-style modulation, highlighting adaptive semantic integration mechanisms that support fusion of diverse priors.
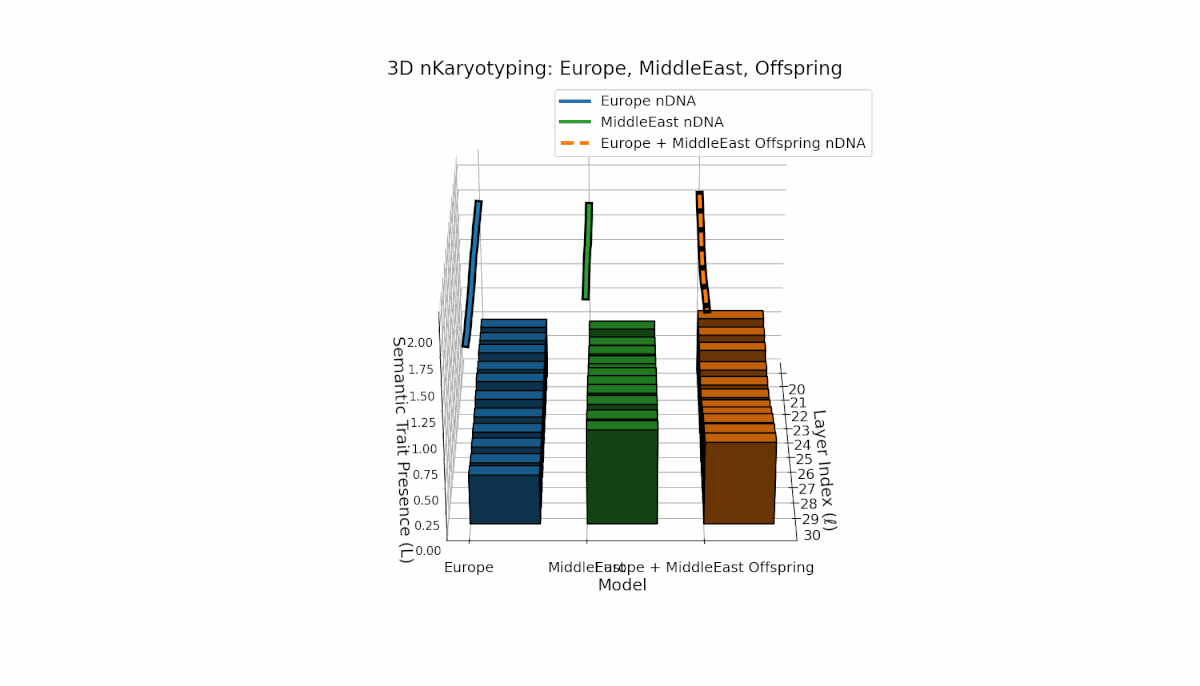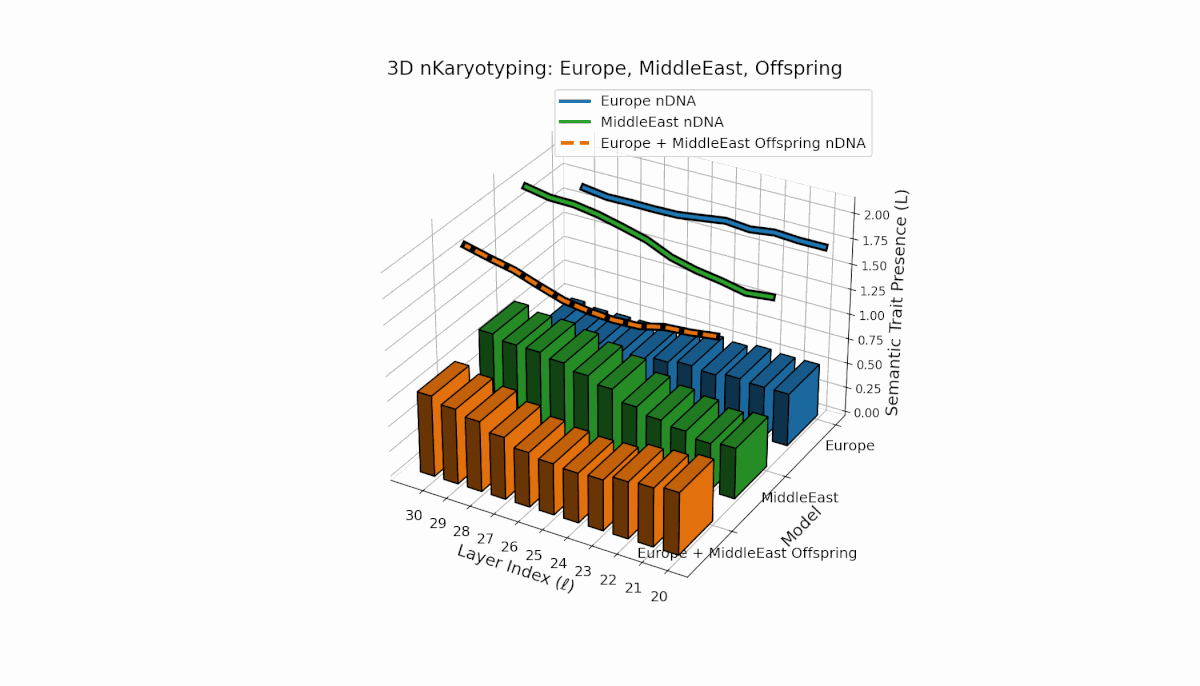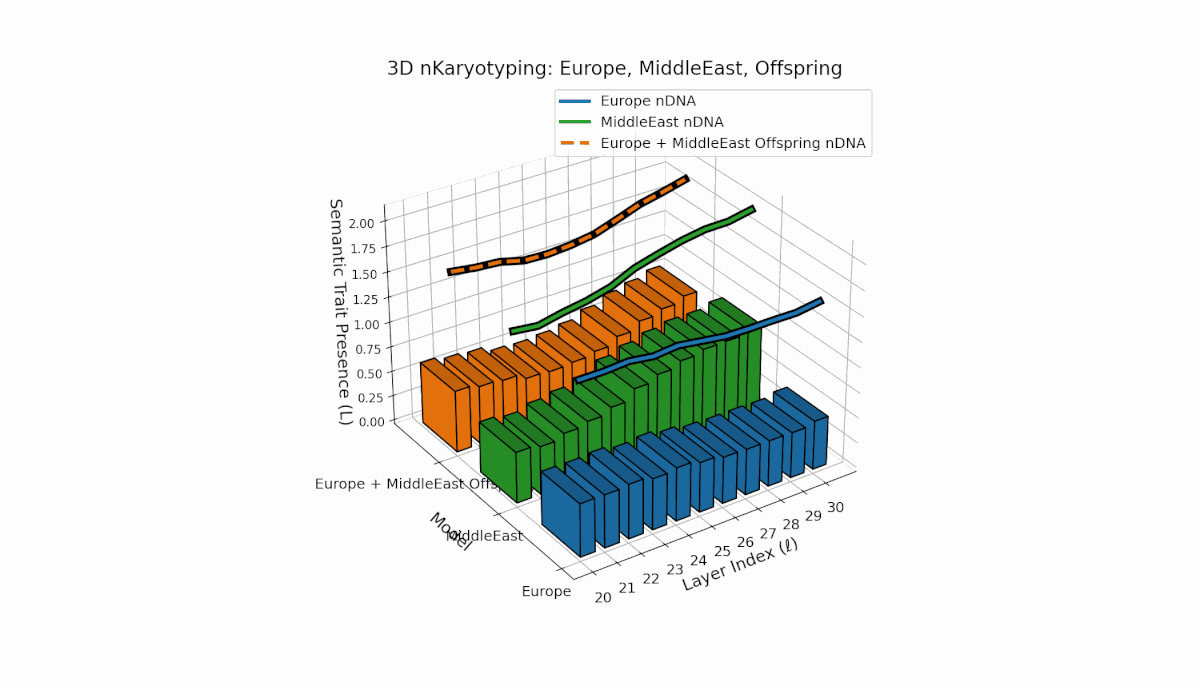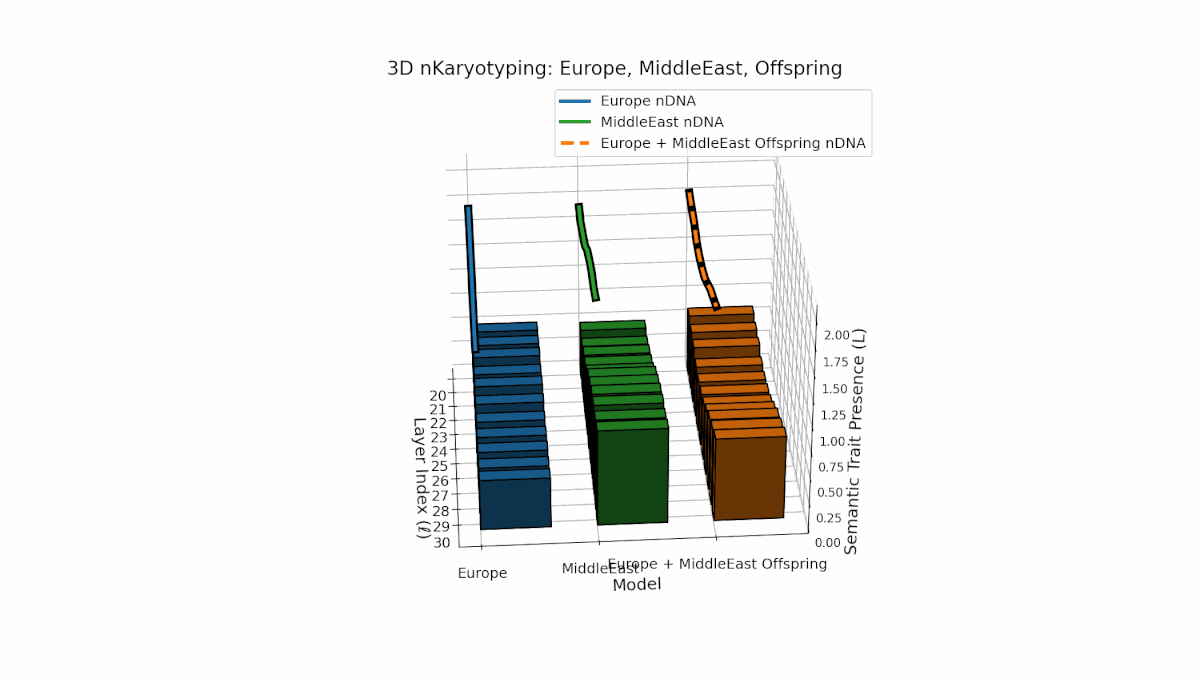
Europe exhibits a broad distribution of semantic chromosomes across early-to-mid layers l = 20–27, with moderate spectral curvature and thermodynamic length indicating stable semantic processing. Middle East traits concentrate in mid layers (l = 25–29) with pronounced curvature peaks, reflecting distinct latent semantic specialization. The offspring shows a hybrid inheritance mosaic with novel traits emerging in late layers and evidence of latent space reconfiguration, revealing nonlinear dynamics and epigenetic-like phenomena in semantic fusion.
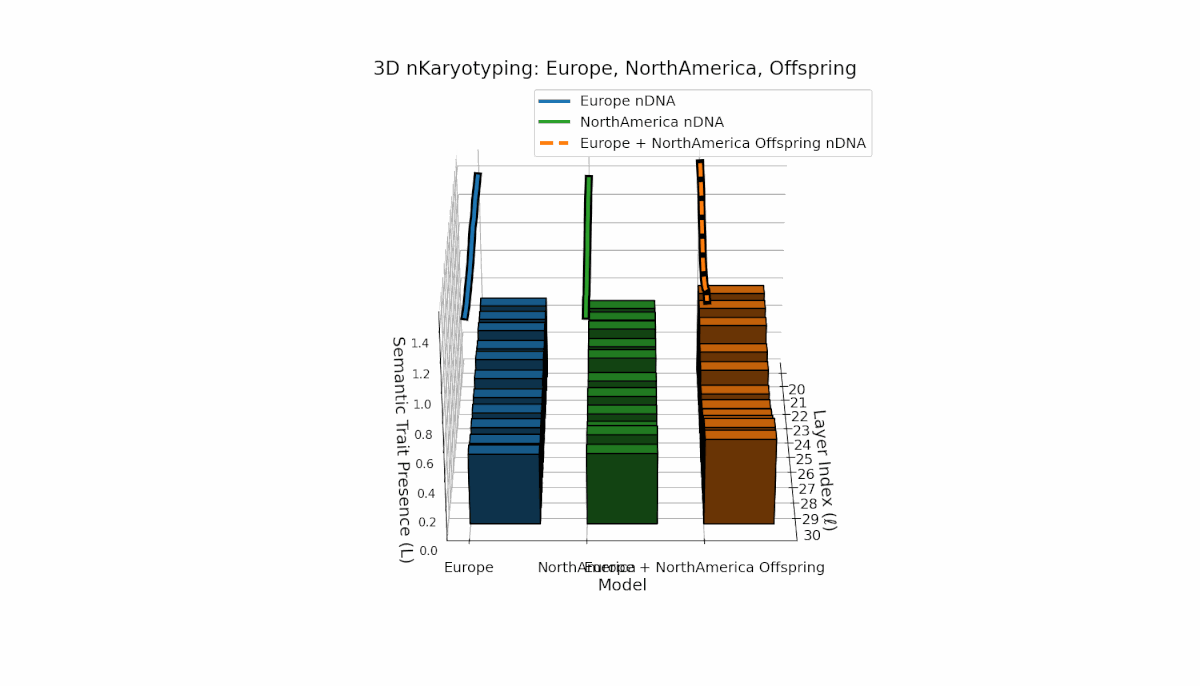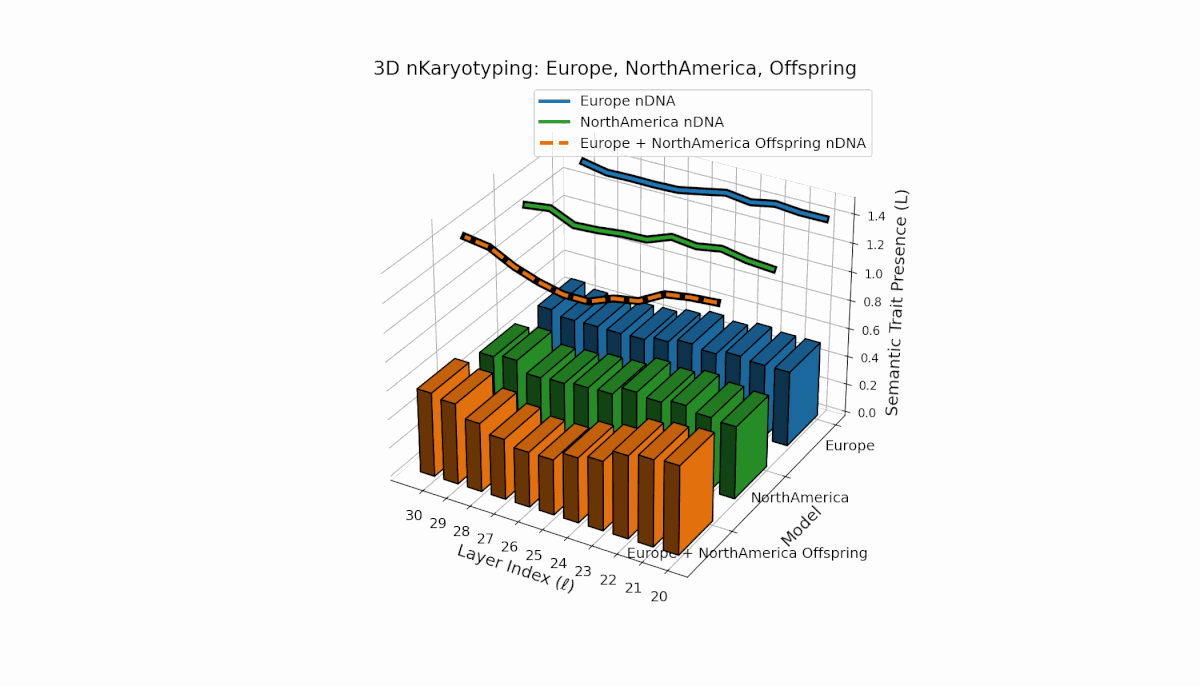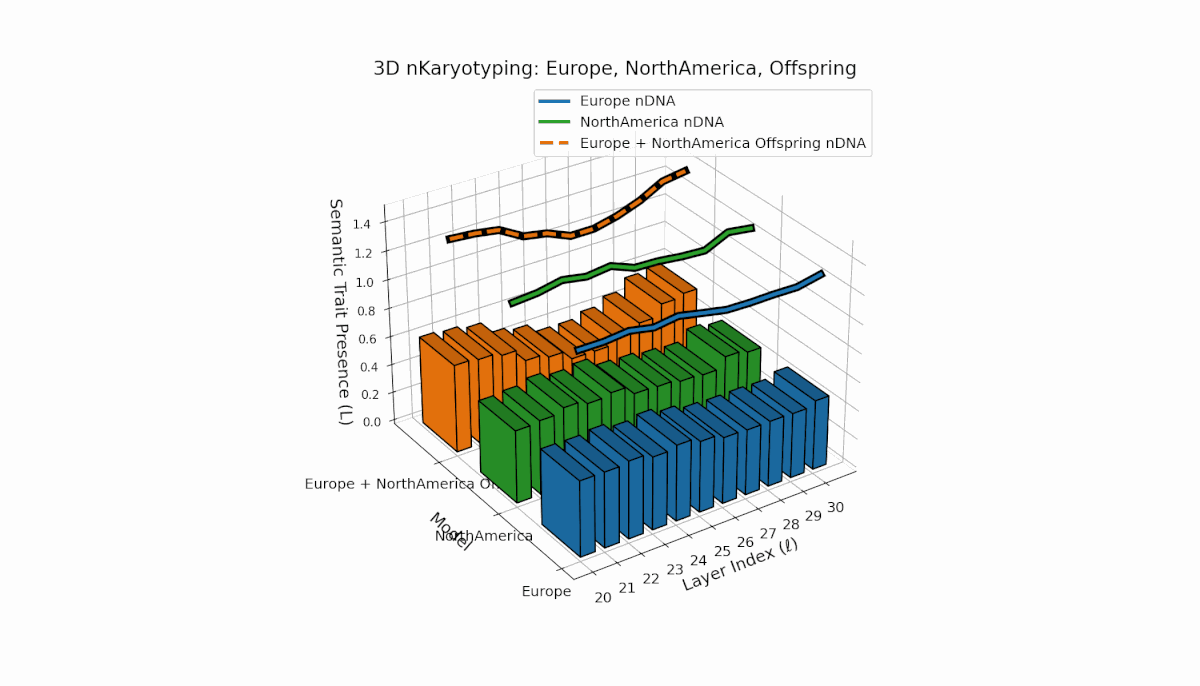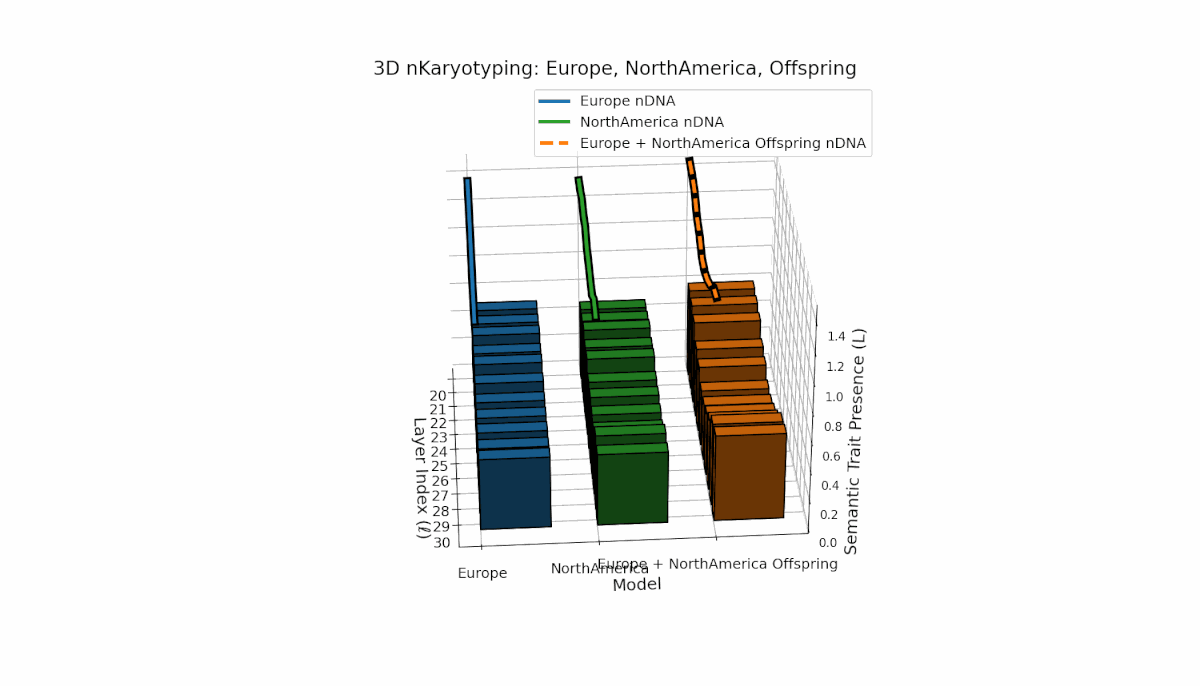
Europe shows a stable presence of semantic chromosomes across layers l = 20–30, with moderate spectral curvature (κl ≈ 0.4–0.6) and consistent thermodynamic length (Ll ≈ 0.6–1.5), indicating robust semantic features. North America exhibits stronger early-to-mid layer trait intensities with spectral curvature roughly (κl ≈ 0.3–0.5) and fluctuating thermodynamic lengths (Ll ≈ 0.4–1.0), revealing a dynamic semantic landscape. The offspring demonstrates complex nonlinear recombination of parental traits, reflecting latent semantic regulatory interplay and emergence of hybrid knowledge representations.
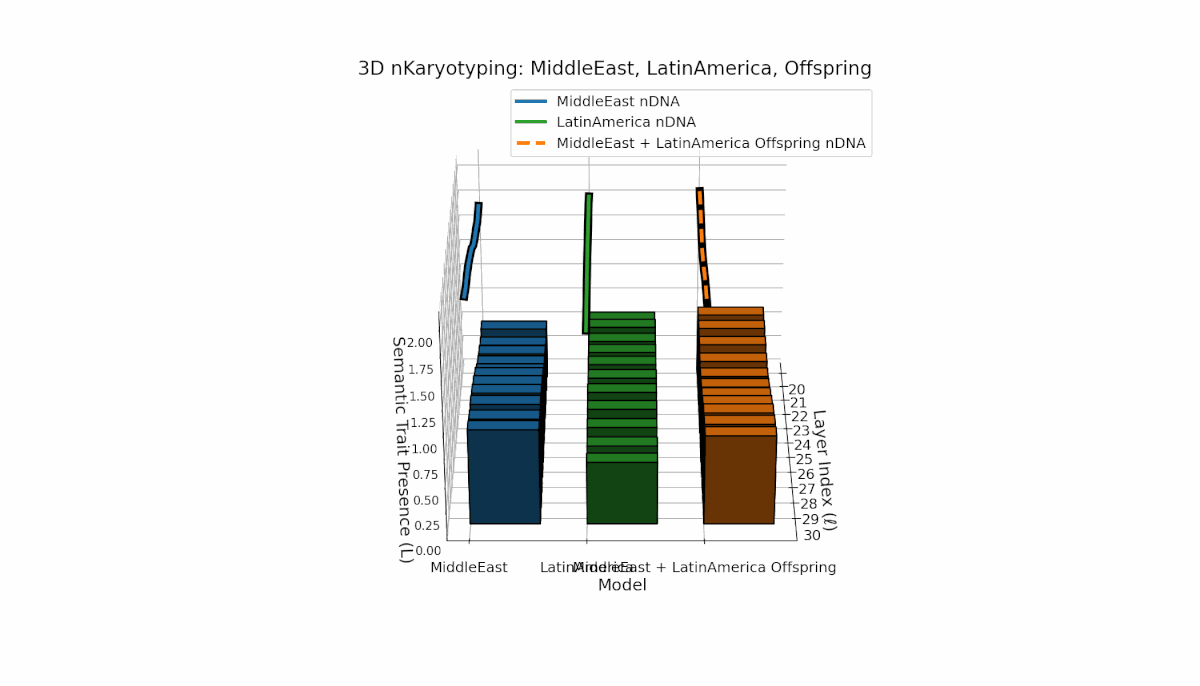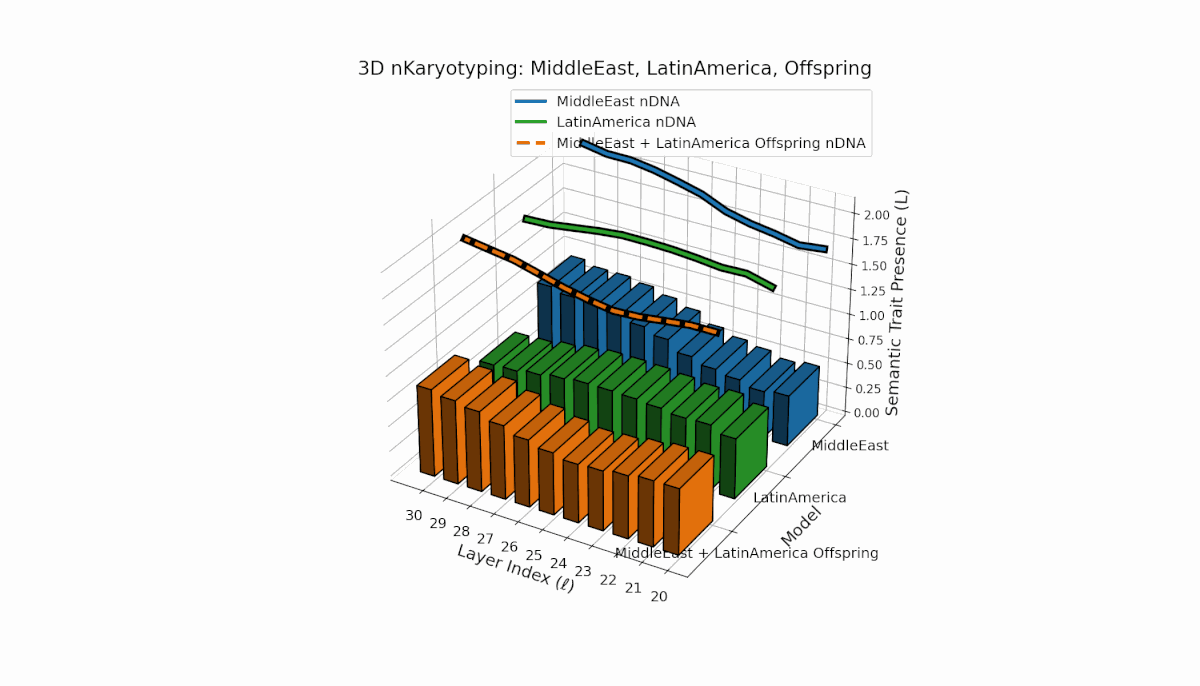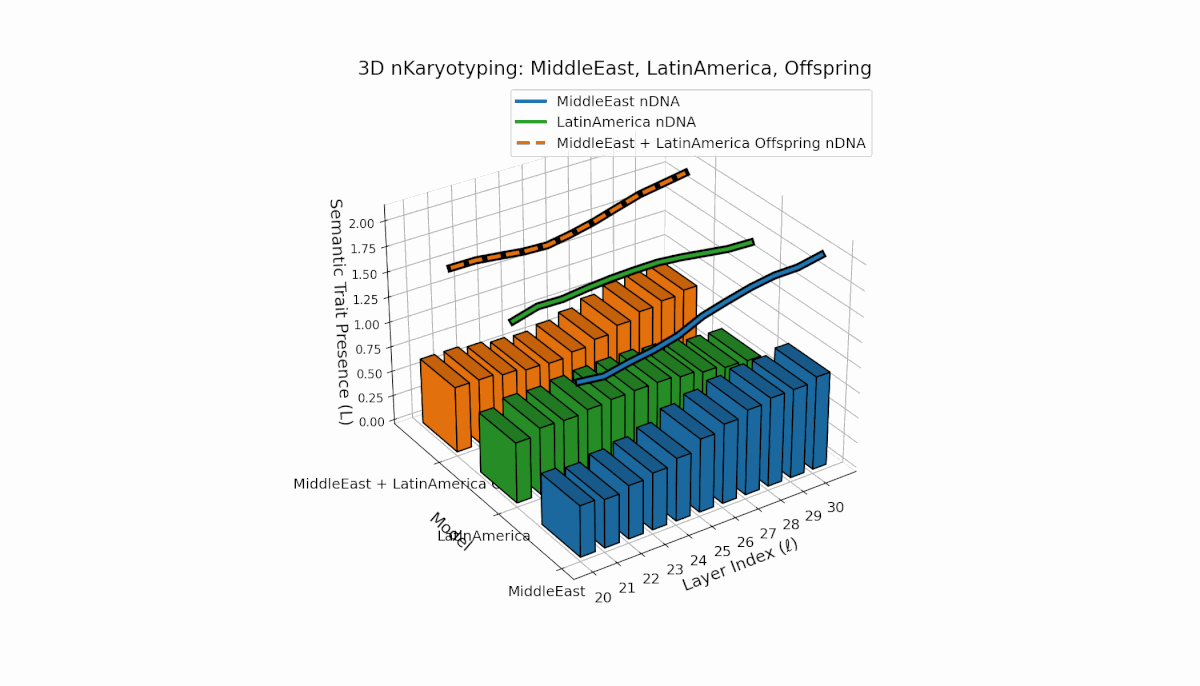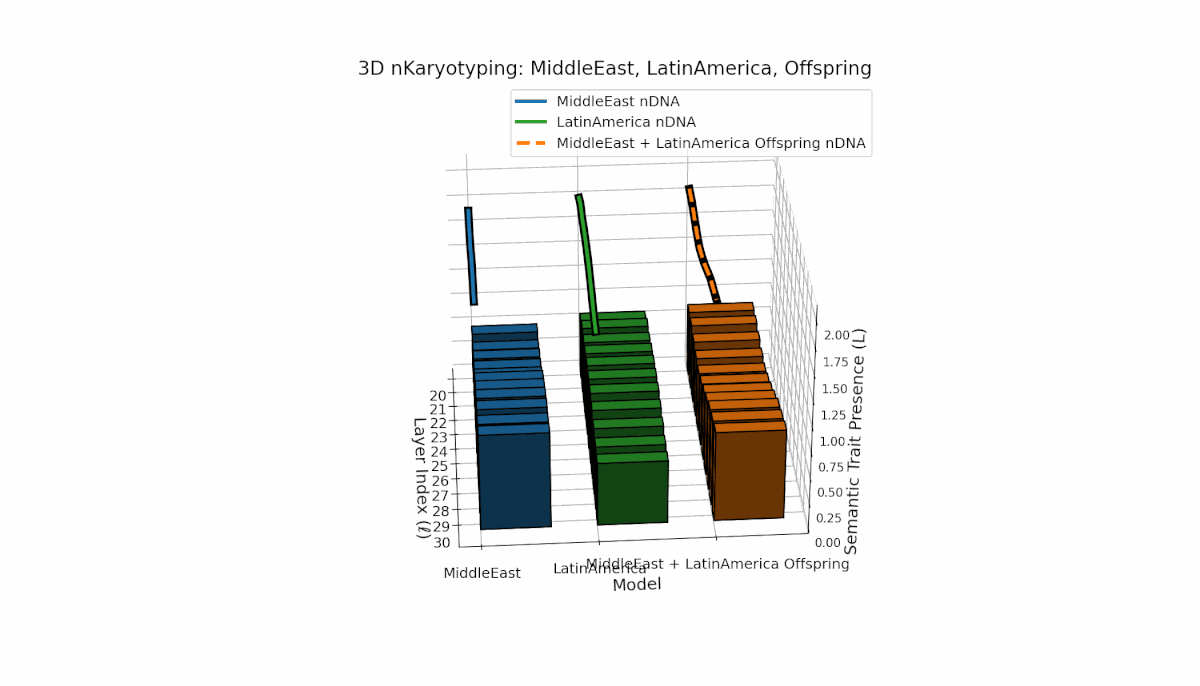
The Middle East model exhibits a pronounced semantic chromosome presence in early layers (l = 20–24) with high spectral curvature (κl ≈ 0.55–0.75) indicating specialized semantic features. Latin America shows a steady increase in semantic chromosomes in deeper layers (l = 25–30) with thermodynamic length rising from Ll ≈ 0.6 to 1.0, reflecting gradual semantic integration. The offspring displays a sophisticated mosaic combining parental traits and novel emergent features primarily in mid-to-late layers, illustrating latent space reorganization and epigenetic-like adaptations.
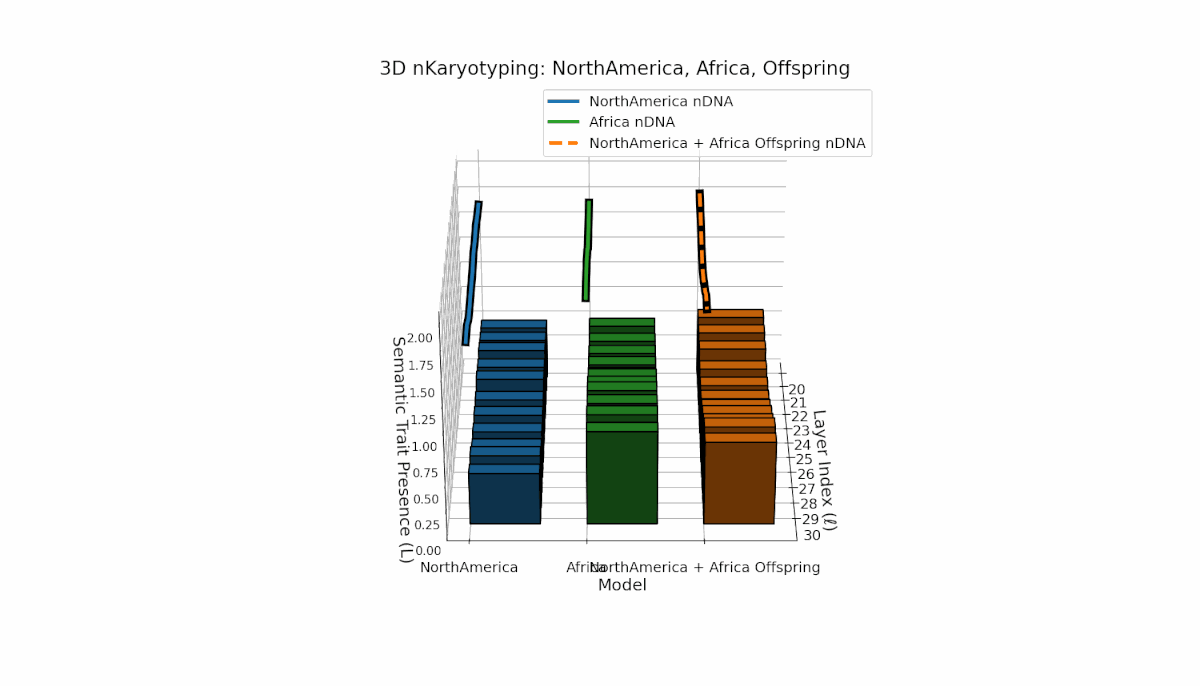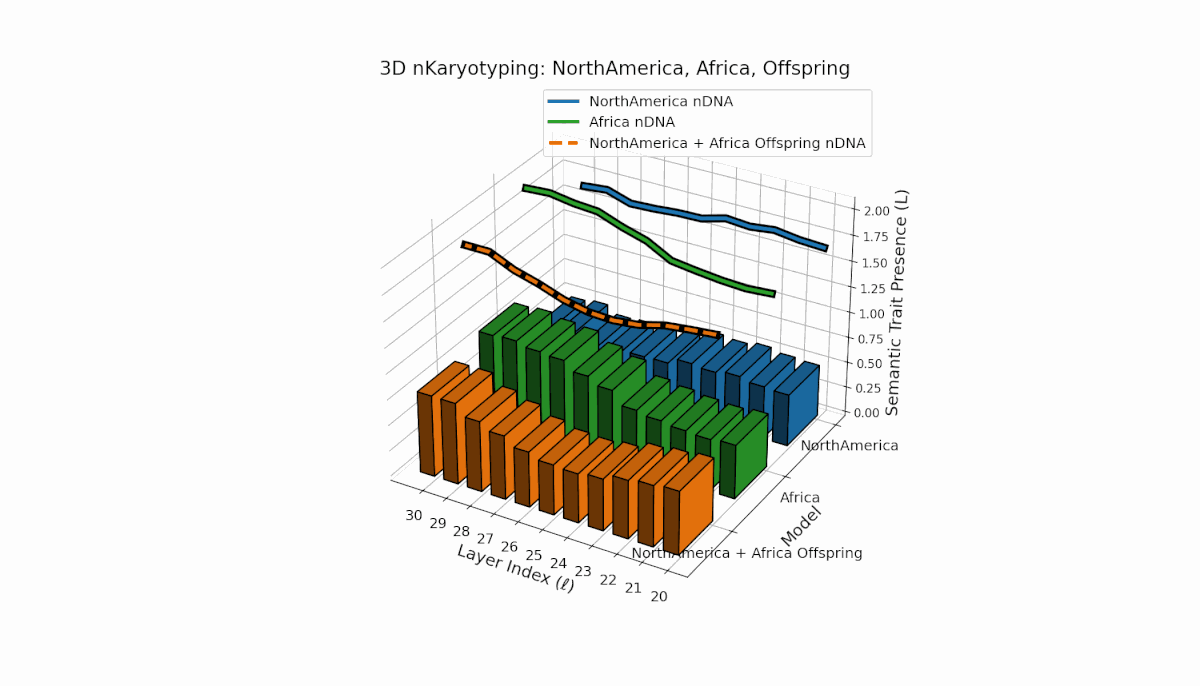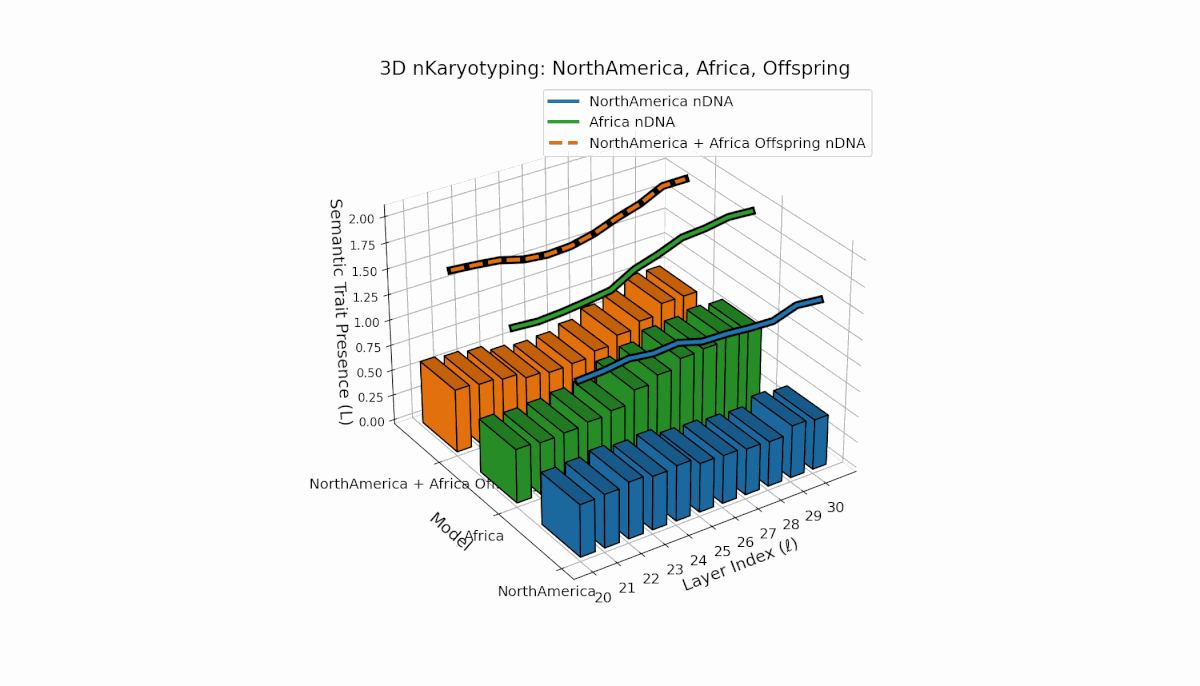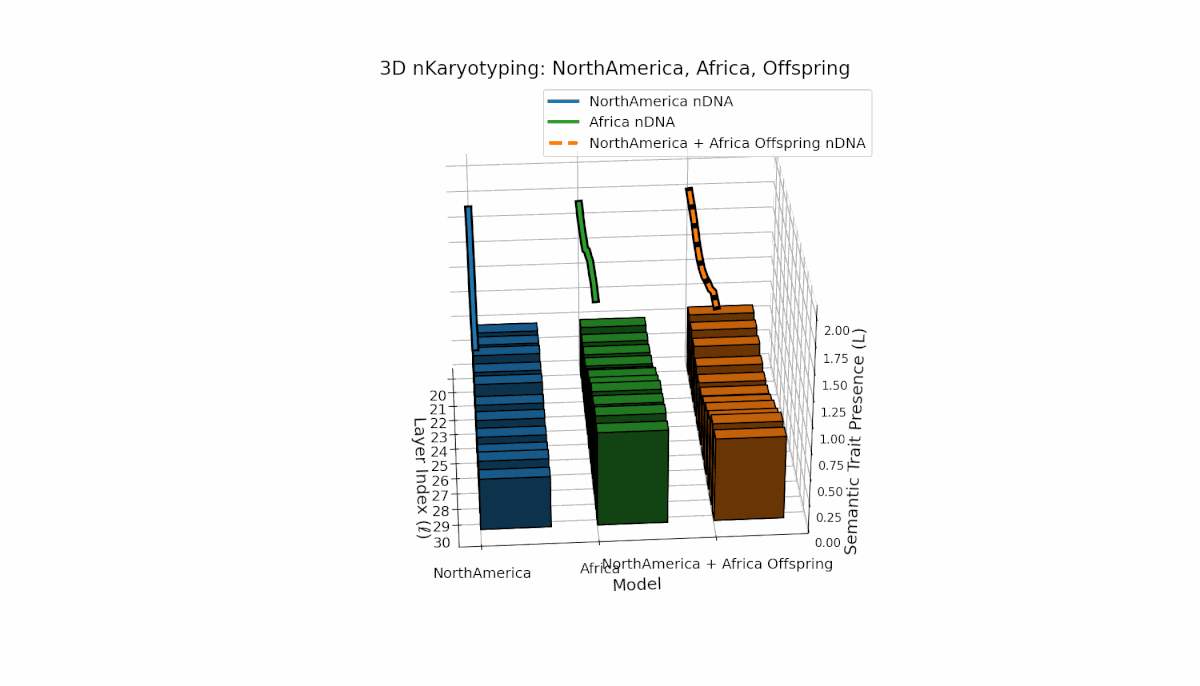
North America exhibits semantic chromosomes primarily in early to mid layers (l = 20–26) with relatively smooth spectral curvature (κl ≈ 0.3–0.5), while Africa shows broader early-layer trait concentration with elevated thermodynamic length (Ll ≈ 0.8–1.2), indicating richer semantic activity. The offspring displays a complex mosaic pattern, merging Africa’s early-layer traits with North America’s mid-layer dominance and featuring novel emergent semantic chromosomes. These patterns suggest intricate semantic regulatory dynamics and epigenetic-like mechanisms guiding semantic fusion and inheritance in model recombination.
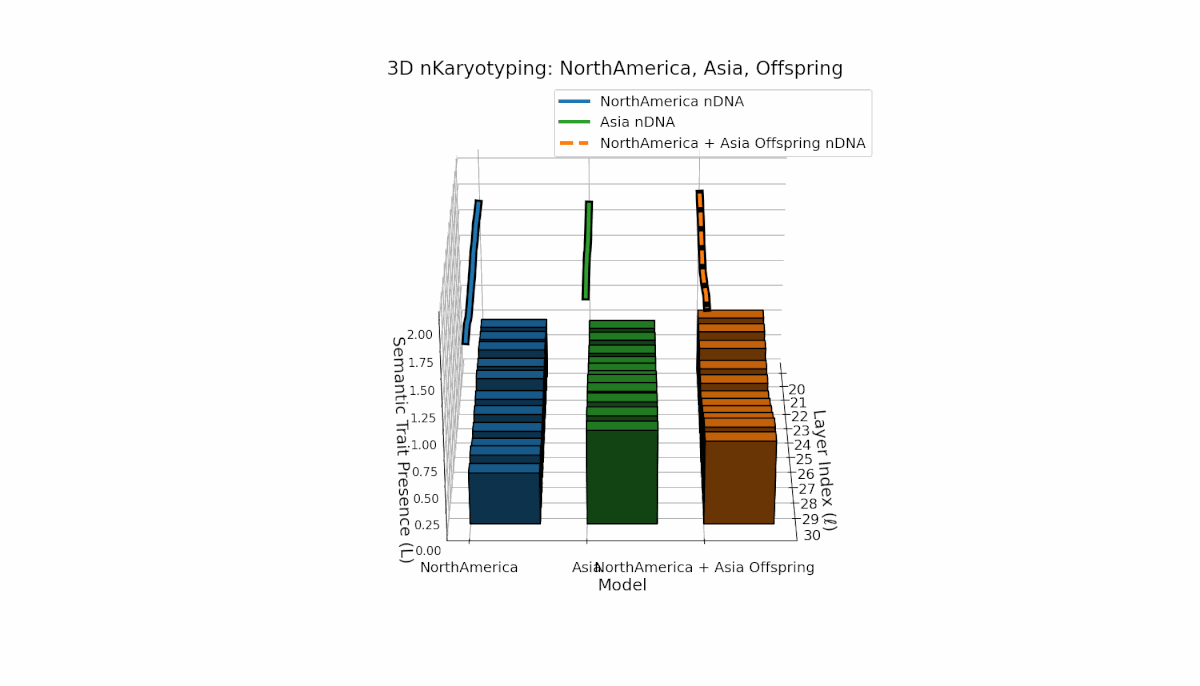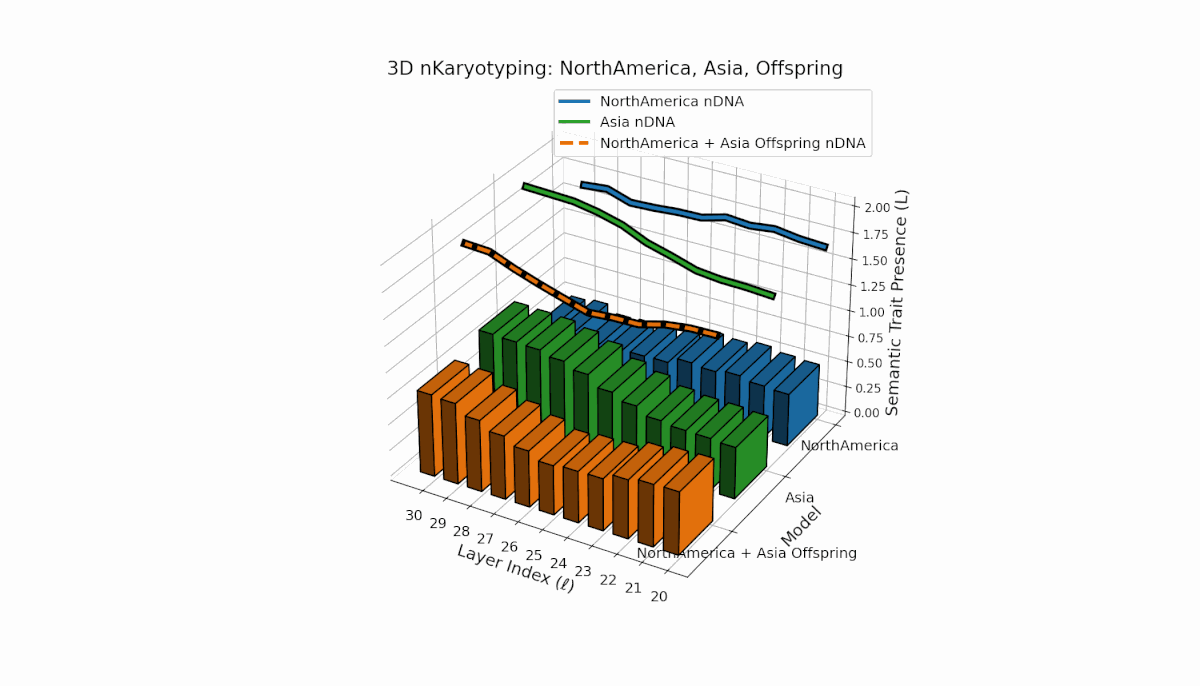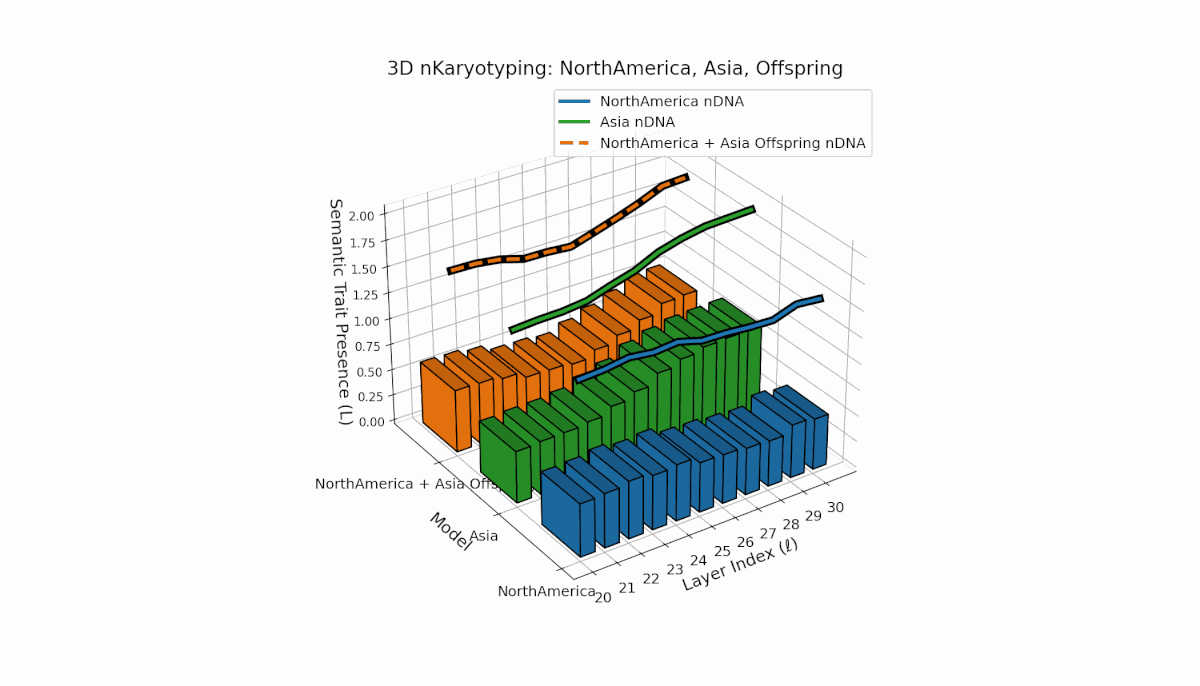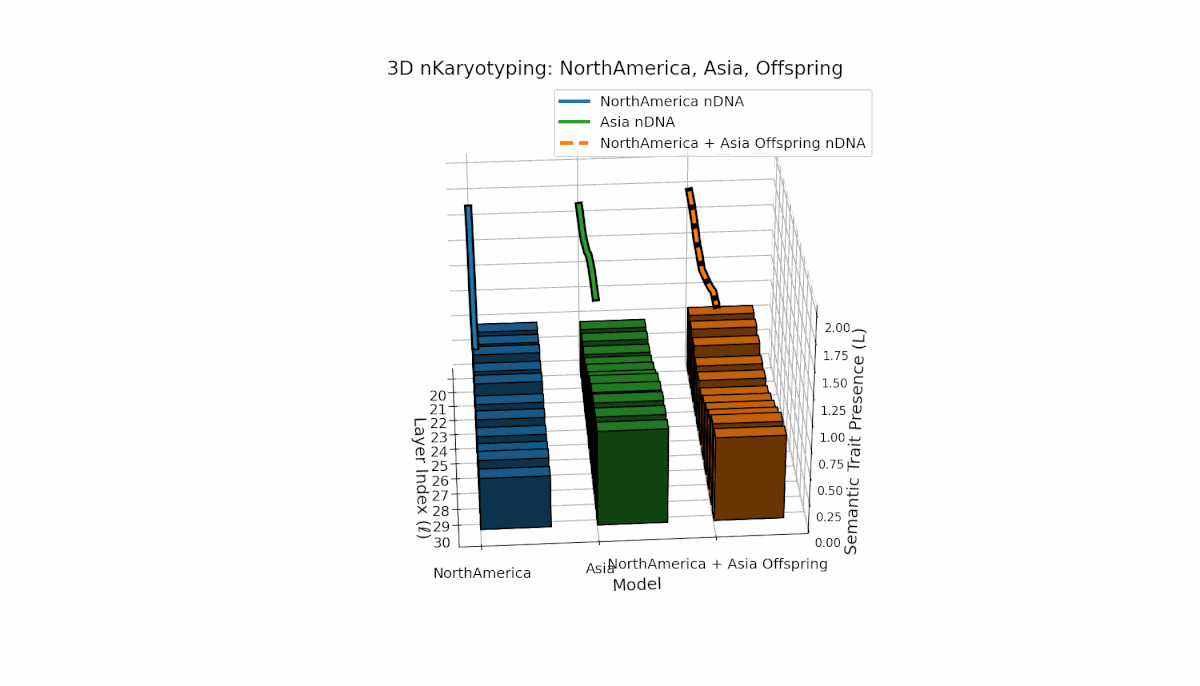
North America shows pronounced early-layer semantic chromosomes across layers l = 20–24, with moderate spectral curvature (κl ≈ 0.35–0.5) and stable thermodynamic length (Ll ≈ 0.3–0.45), reflecting strong early semantic imprinting. Asia exhibits sustained deep-layer traits from l = 25–30, with increased thermodynamic length (Ll ≈ 0.4–0.6), indicating prolonged semantic integration. The offspring demonstrates layerwise recombination and novel trait emergence, revealing nonlinear epigenetic-like modulation and semantic plasticity analogous to biological gene regulatory networks driving phenotypic diversity.
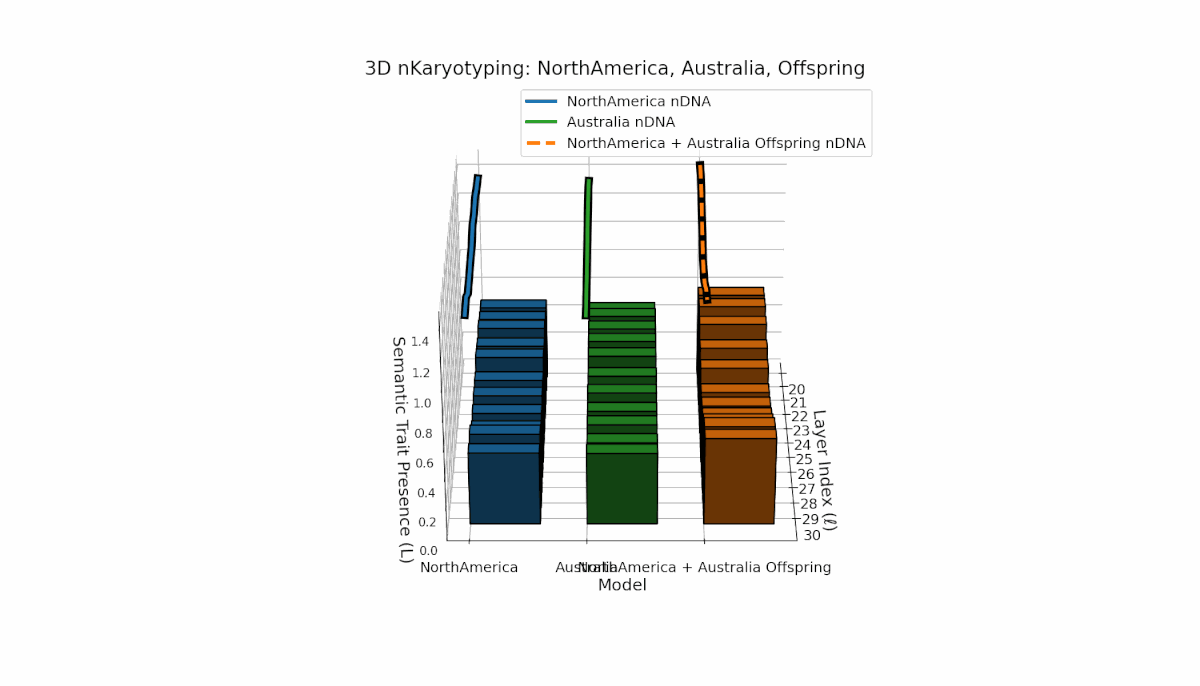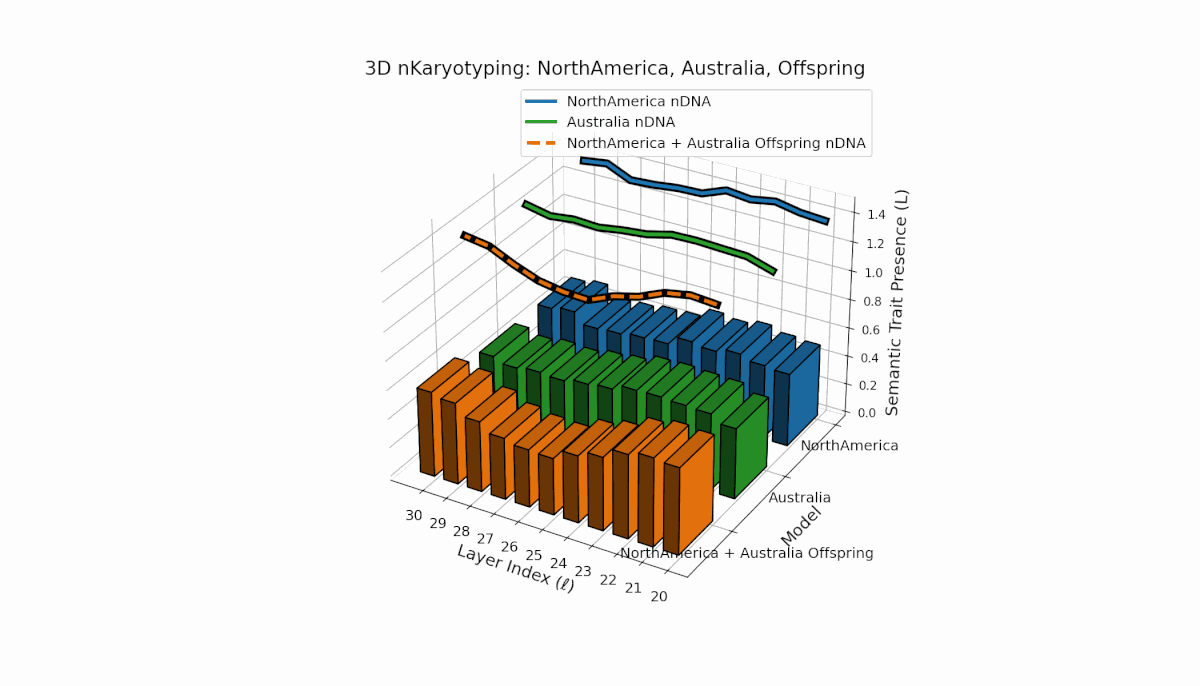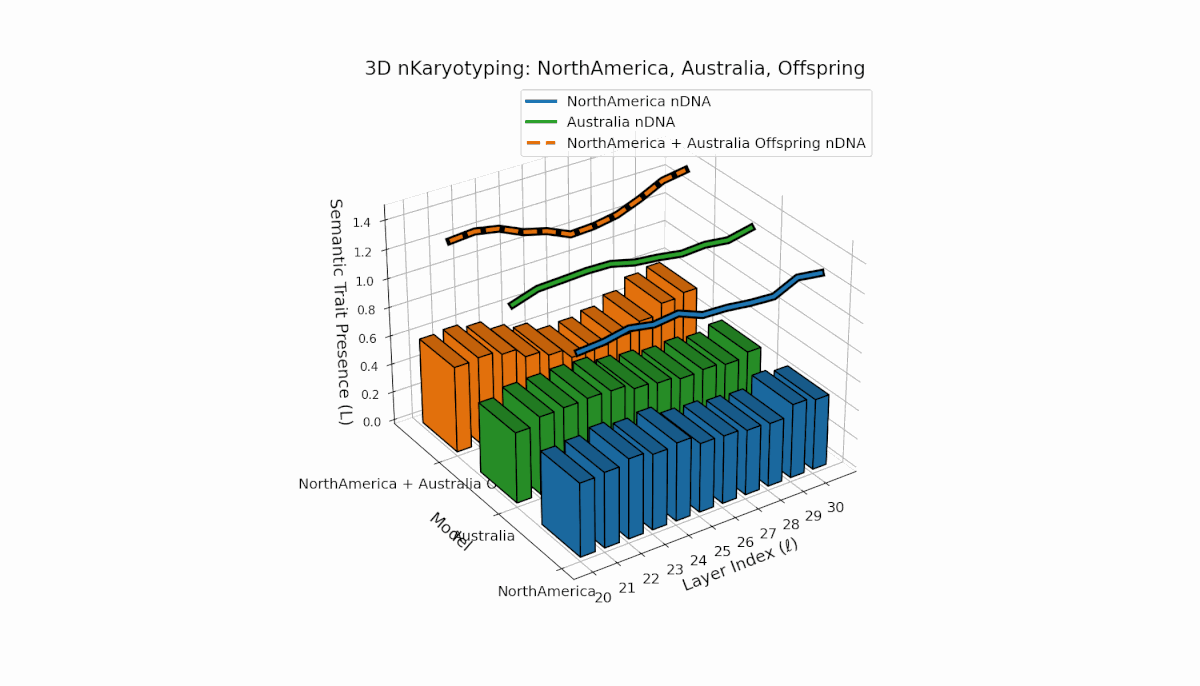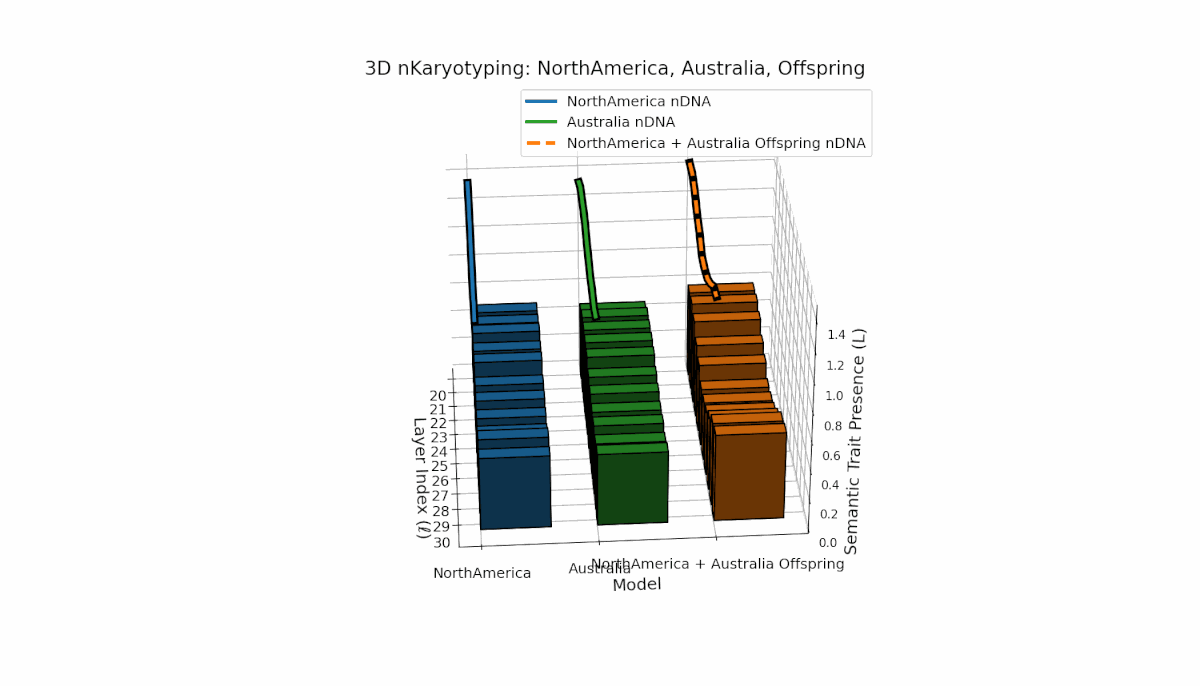
North America shows pronounced semantic chromosomes from layers l = 20–26 with distinct oscillations in trait expression and thermodynamic length. Australia exhibits balanced trait distribution across layers l = 20–30 with moderate spectral curvature. The offspring demonstrates complex recombination and layered trait blending, reflecting latent semantic plasticity and integrative dynamics in large model fusion.
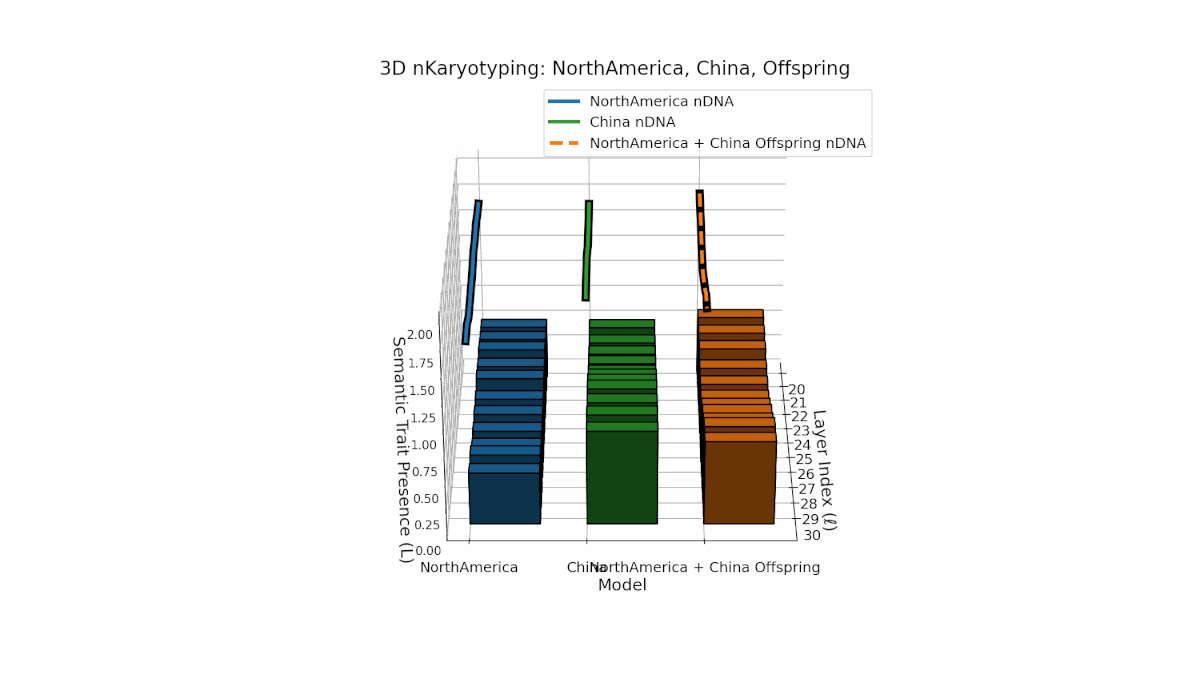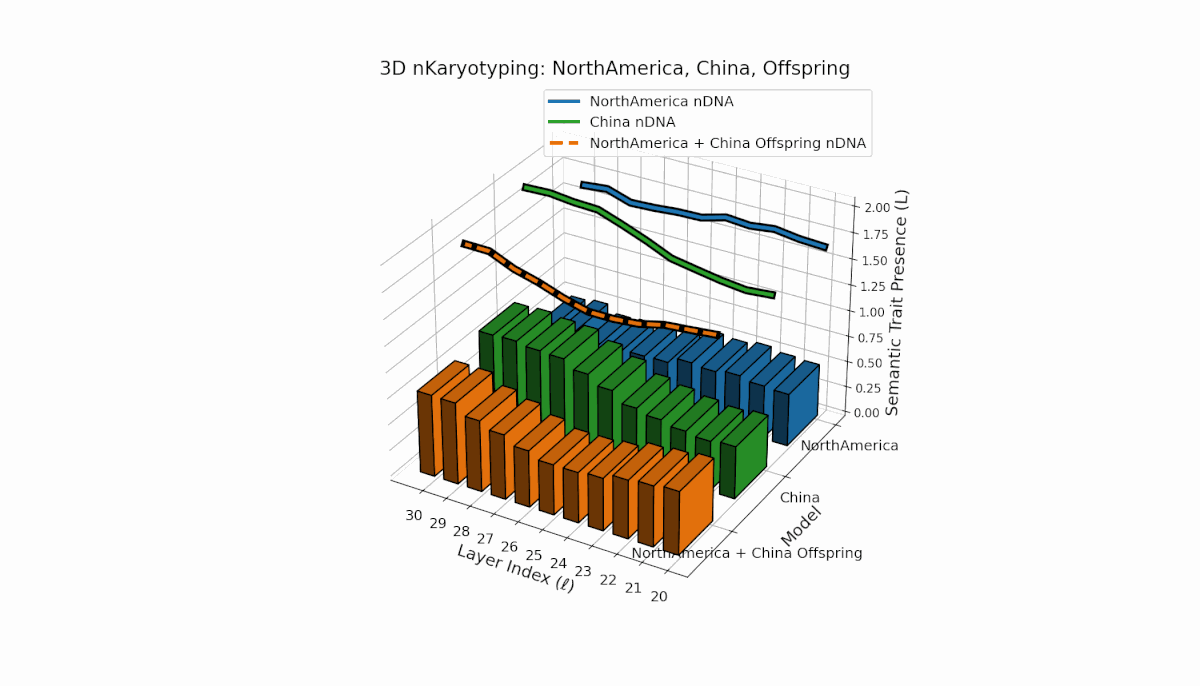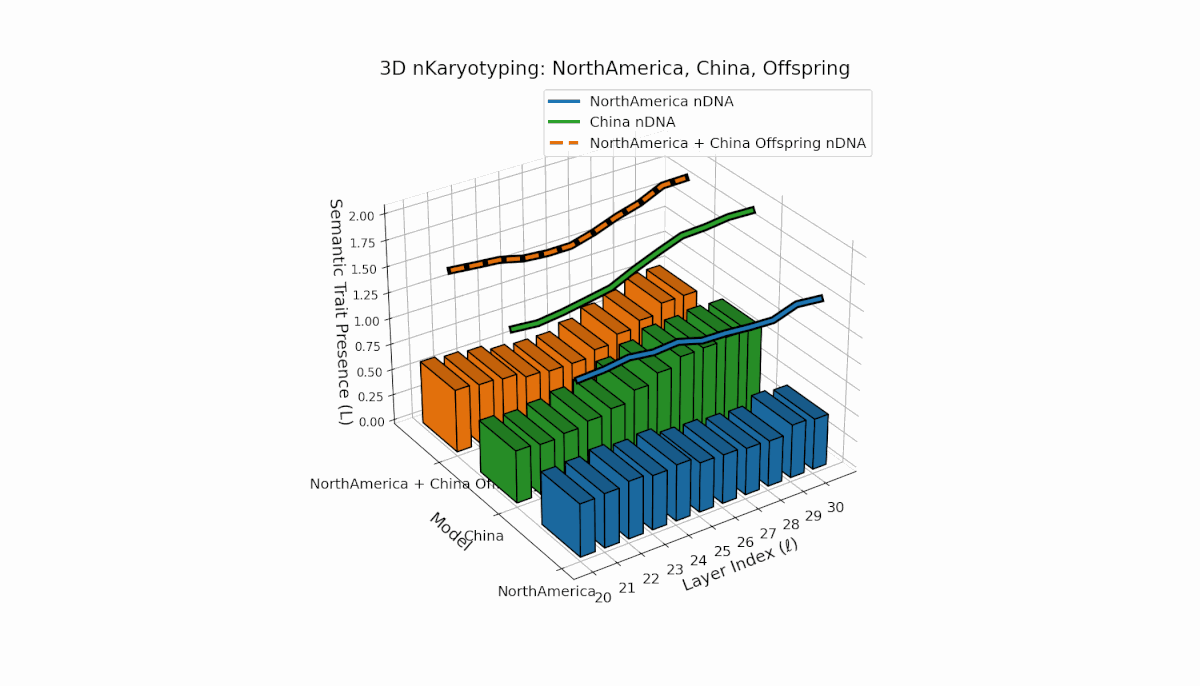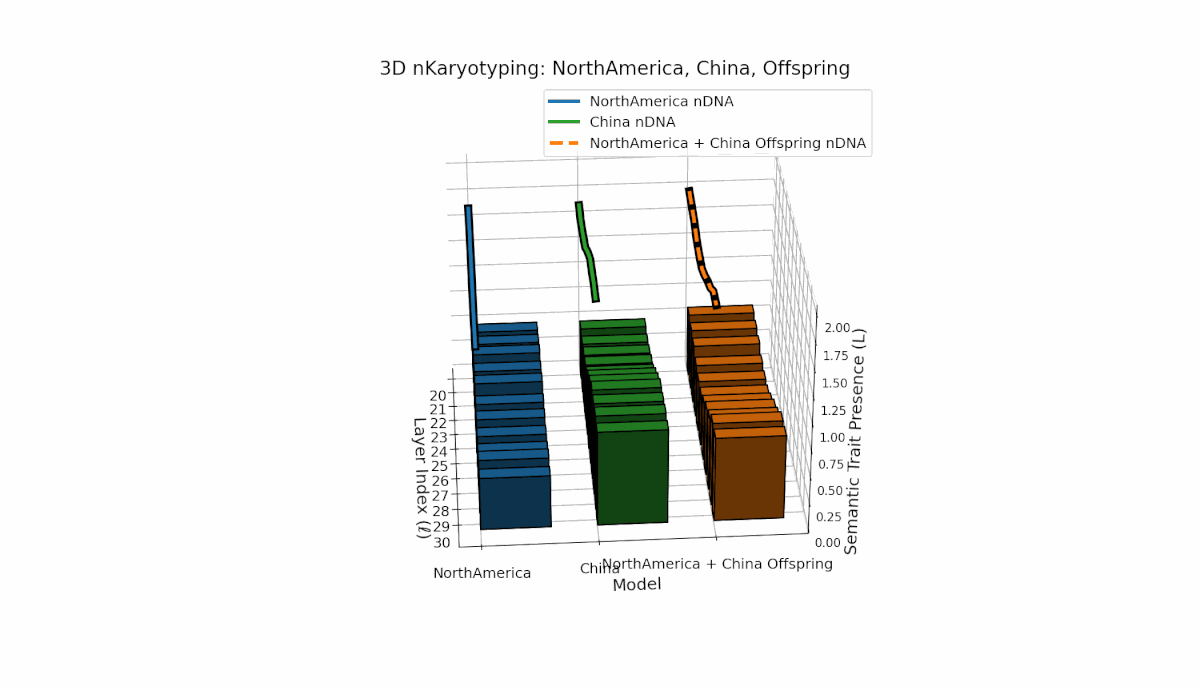
North America shows early-layer dominance with pronounced trait fluctuations across layers l = 20–26, while China exhibits sustained deep-layer activation and elevated thermodynamic length from l = 25–30. The offspring combines these contrasting patterns, revealing nuanced latent space trajectories that reflect cross-cultural semantic fusion and suggest emergent representational features beyond additive inheritance.
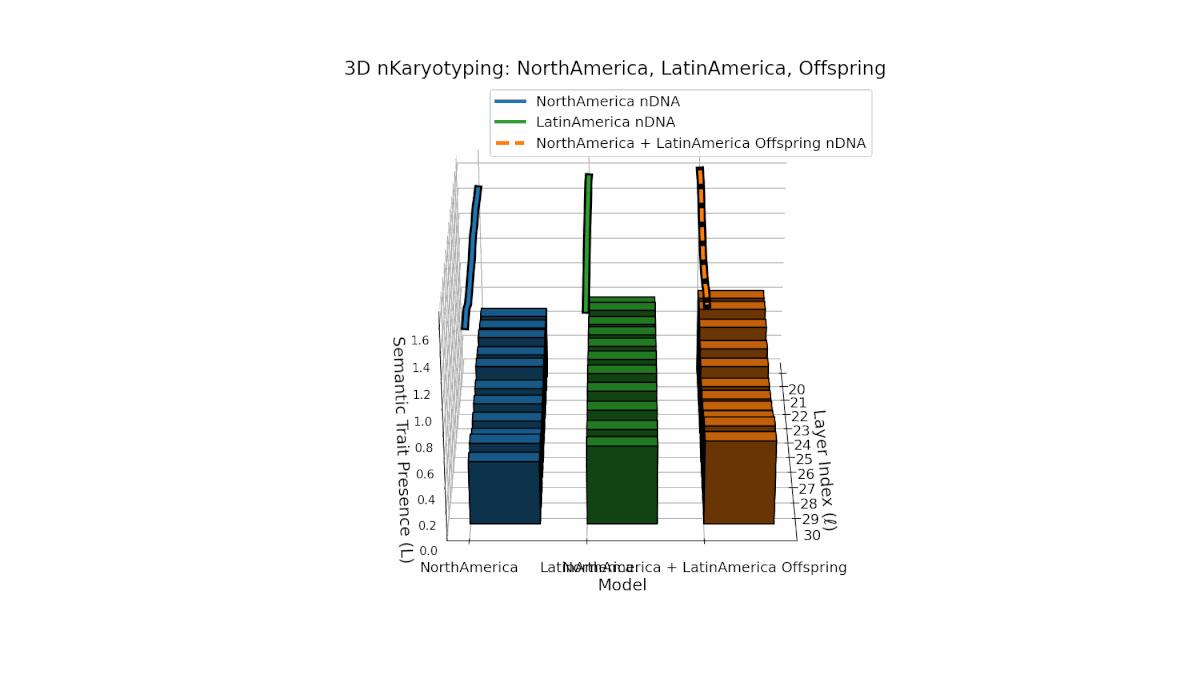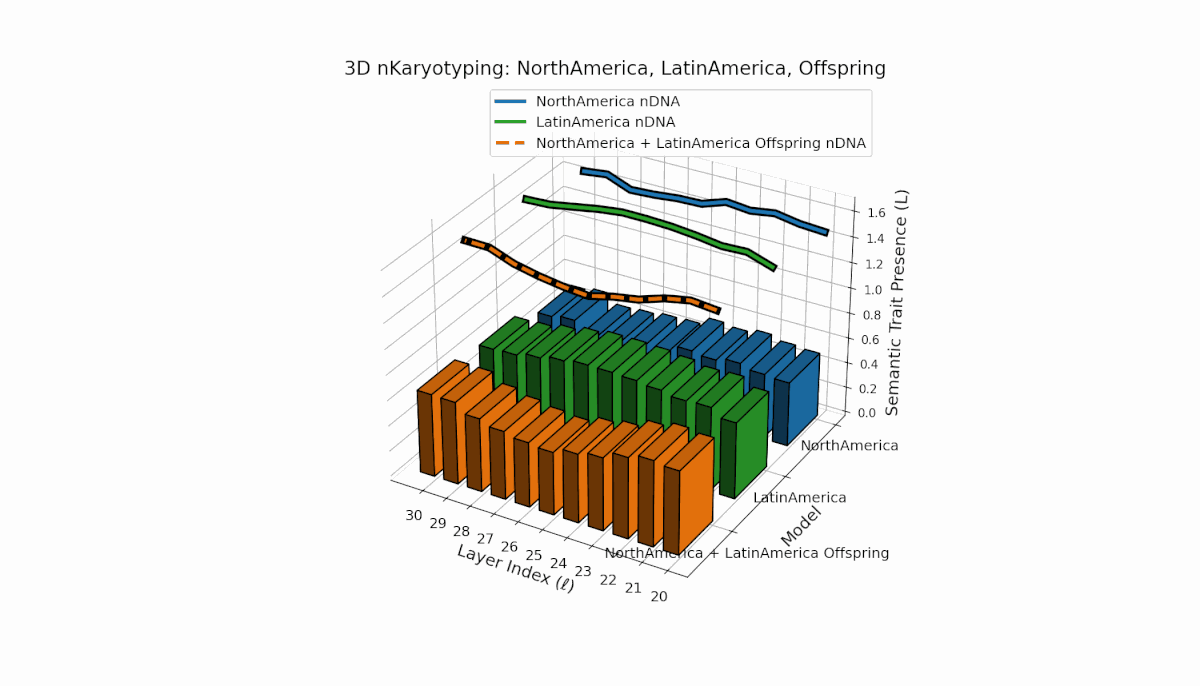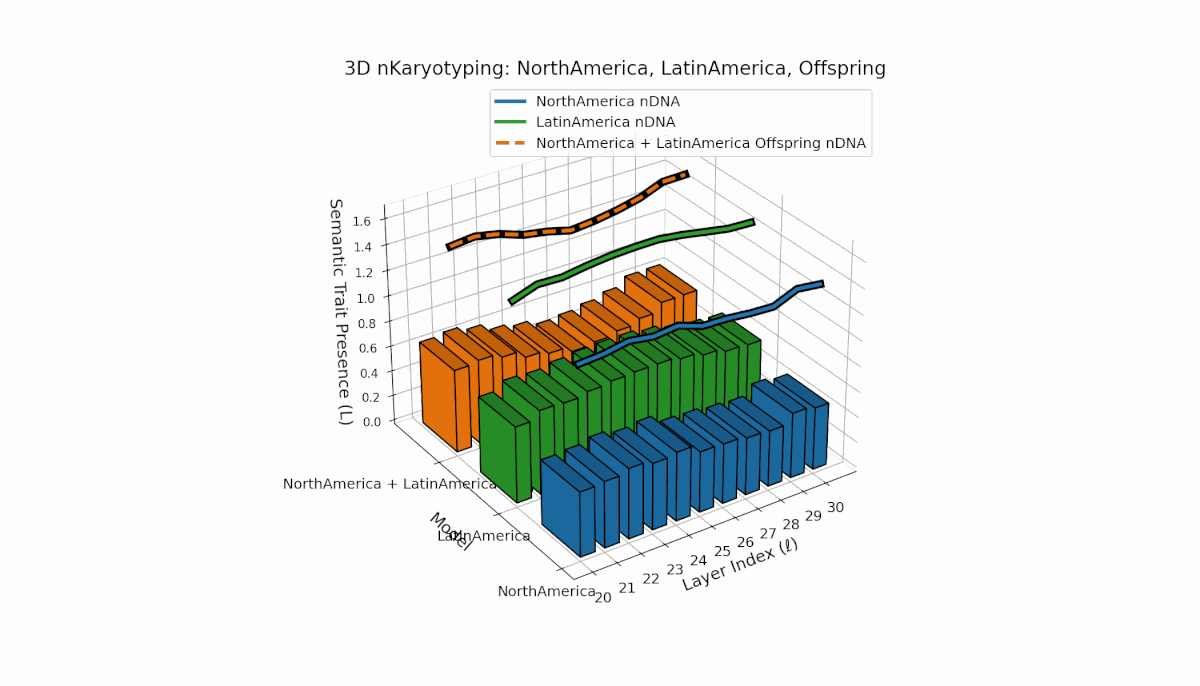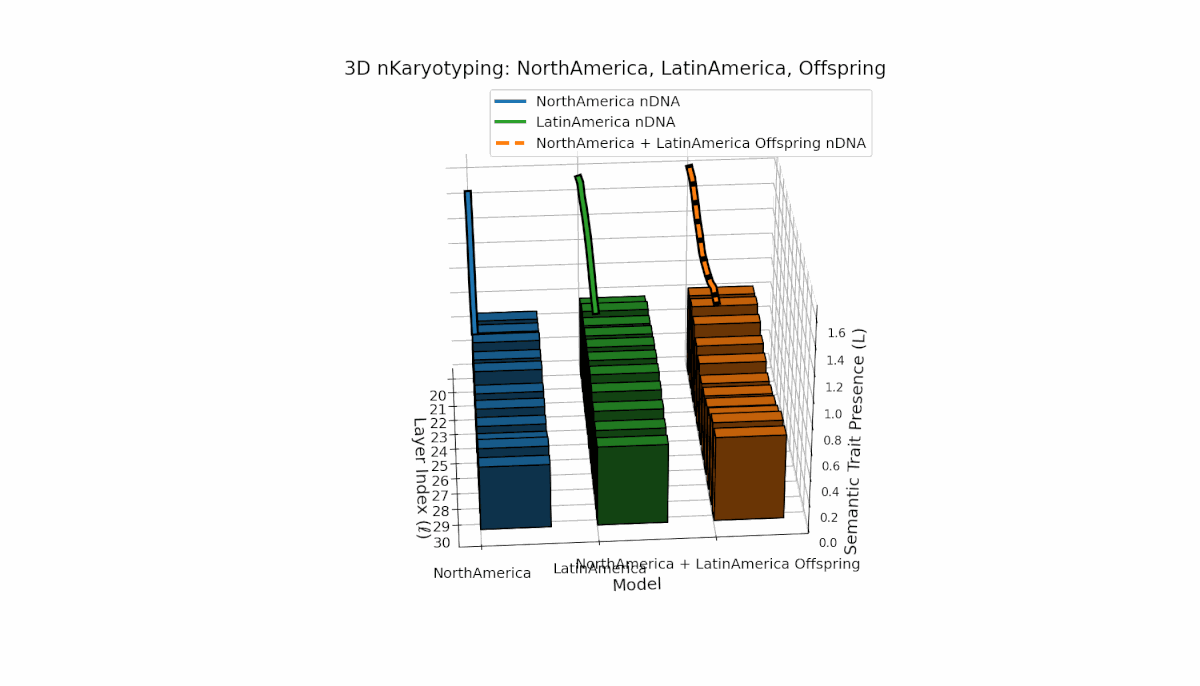
North America shows layered semantic chromosome presence primarily in early to mid layers (l = 20–26), while Latin America exhibits a steady increase in trait presence in later layers (l = 25–30). Their offspring reveals a hybrid latent profile combining early and late traits, evidencing emergent semantic structures and dynamic recombination reflective of complex cultural-semantic interactions in the nDNA.
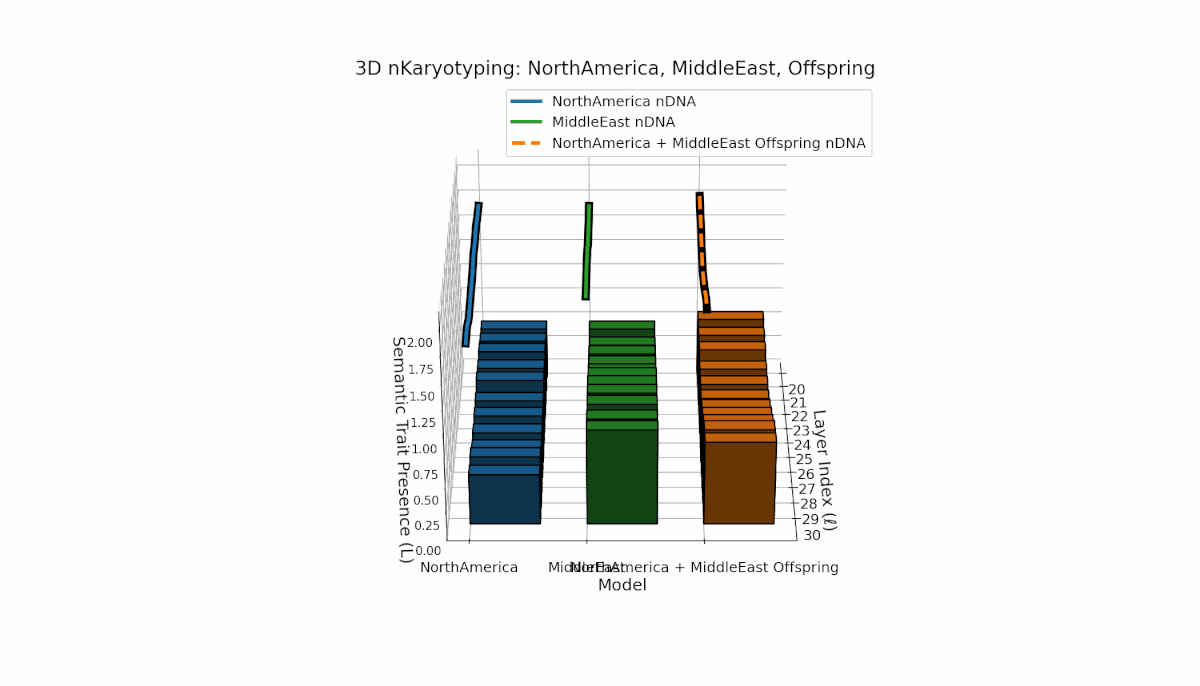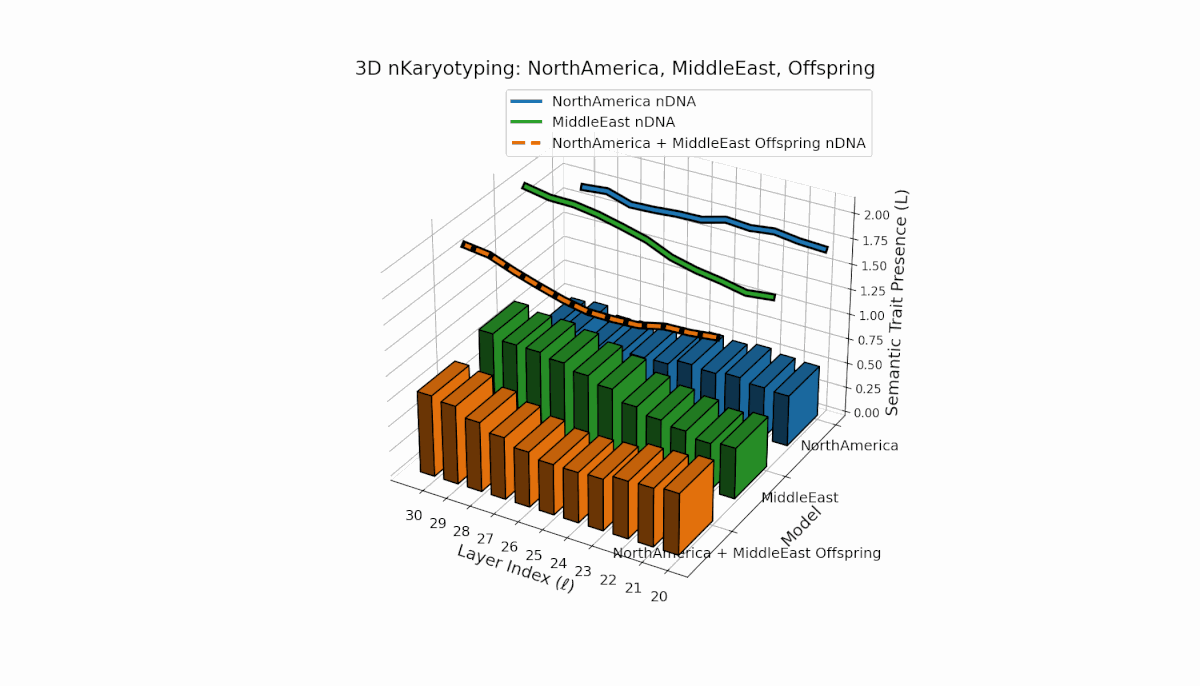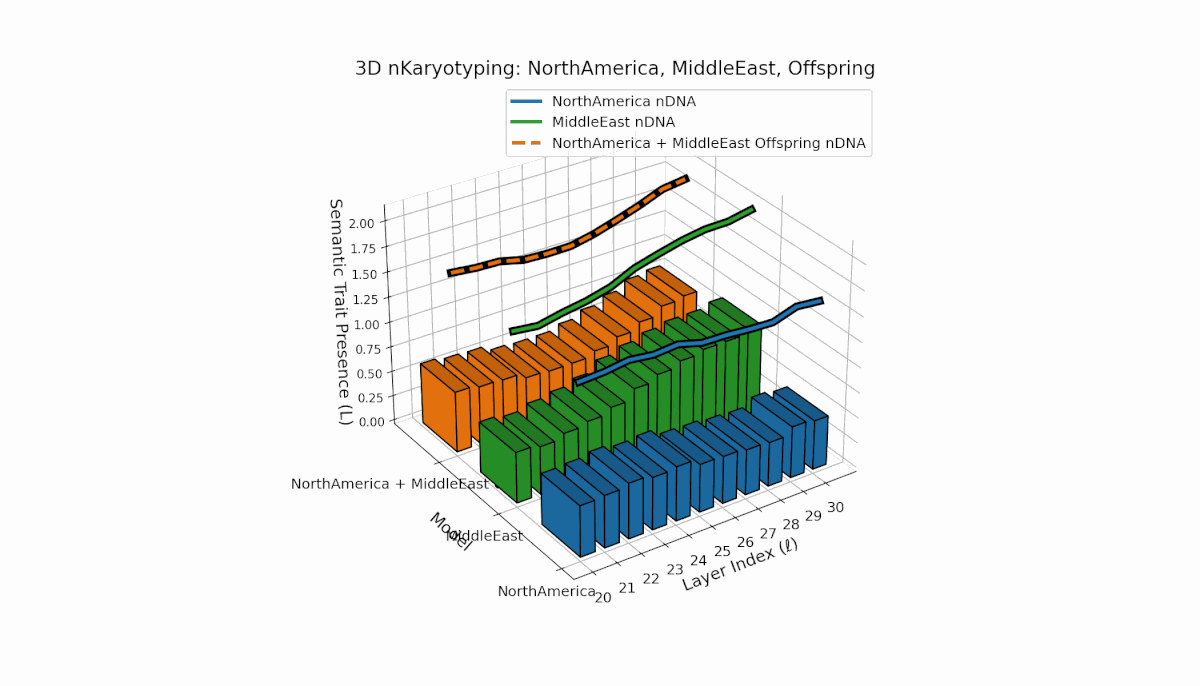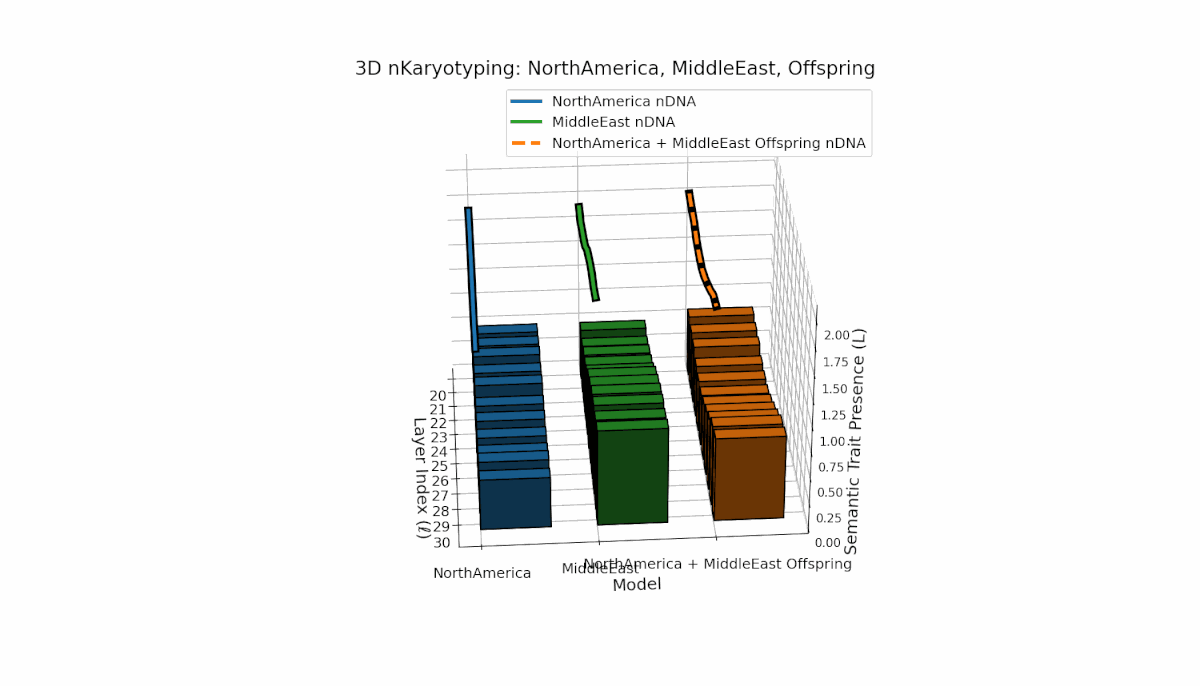
North America exhibits strong early-layer semantic chromosomes with distinct peaks (l = 20–24), while Middle East shows broader mid-to-late layer activation with elevated spectral curvature (l = 25–30). The offspring displays a sophisticated blend of layer-specific recombinations and novel semantic patterns, reflecting complex latent space reshaping akin to biological epigenetic modulation.
This comprehensive visualization presents the layered semantic geometry of cultural fusion models through the lens of nKaryotyping, a diagnostic framework quantifying trait presence $L(\ell)$ and spectral curvature $\kappa(\ell)$ across 28 culture pair offspring models. The figure displays three key dimensions: (1) Bar plots (bottom layers): Represent semantic trait presence $L(\ell)$ across transformer layers $\ell = 20\ldots30$ for each parent culture and their offspring, with heights ranging approximately $0.3$–$1.2$ indicating activation intensity. (2) Thick black-edged colored lines: Smooth layerwise trait presence curves $L(\ell)$, providing continuous summaries for each model — two parents (solid blue and green) and the offspring (dashed orange) — highlighting preservation and blending of semantic features across depth. (3) Dotted lines (top): Spectral curvature trajectories $\kappa(\ell)$, reflecting geometric complexity and nonlinear representational structure in embedding space, ranging roughly $0.3$–$0.9$ with oscillations indicating representational bending or epistemic transitions. Layout: The 28 offspring models correspond to all pairwise combinations of eight canonical styles — Europe, North America, Australia, Africa, Asia, China, Middle East, and Latin America — organized along the culture pair index axis, while the layer index axis spans $\ell = 20\ldots30$; explicit spacing between subplots enhances visual clarity; the vertical axis denotes trait and curvature values. Key insights: (1) Heterogeneous Semantic Inheritance: Offspring profiles blend parental features with nuanced deviations; intermediate layers often show enhanced or attenuated activation, evidencing nonlinear fusion dynamics. (2) Spectral Curvature as Epistemic Indicator: Peaks and troughs in $\kappa(\ell)$ identify layers of sharp representational bending, potentially correlating with functional transitions or emergent semantic properties distinct from either parent. (3) Cultural Signature Distinctions: Parent cultures show distinct curvature and trait patterns — e.g., Asia and Middle East exhibit pronounced oscillatory curvature, while Europe and North America maintain more stable, literal semantic structures. (4) Offspring Diversity: The spectrum of curvature and trait presence across offspring reflects rich generalization/specialization, informing targeted fine-tuning for culturally-aware AI. An accompanying interactive 3D visualization enables detailed exploration of each offspring’s layered semantic topology and curvature dynamics.
References
[1] Steven H. Strogatz “Nonlinear Dynamics and Chaos: With Applications to Physics, Biology, Chemistry, and Engineering” Comprehensive textbook on dynamical systems and bifurcation theory (2018).
[2] Lygeros, John, Deligeorgiou, Panayiotis, and others “Karyotype analysis: chromosome banding and FISH” Methods in Molecular Biology (2019).
[3] Weisenfeld, Neil I and Shendure, Jay “Cytogenetics: A review of techniques” Journal of Medical Genetics (2006).
[4] Fedoriw, Andrei and O’Hagan, Hannah M “Epigenetic regulation of chromatin structure and function in health and disease” Nature Reviews Genetics (2022).
[5] Landry, Christian R, Hartl, Daniel L, and others “Genetic properties influencing the evolvability of gene expression” Science (2007).
[6] Shapiro, Edward, Biezuner, Tamar, and others “Stem cell lineage tracing: techniques, applications and challenges” Nature Reviews Genetics (2013).
[7] Ng, Andrew Y, Jordan, Michael I, and others “On spectral clustering: Analysis and an algorithm” Advances in Neural Information Processing Systems (2002).
[8] Gersen, Steven L and Keagle, Martha L “Clinical Cytogenetics” arXiv preprint (2012).
[9] Leder, Peter and Goldman, Robert “Cytogenetics and Cell Genetics” arXiv preprint (2005).
[10] Mitelman, Felix “Atlas of Chromosome Aberrations in Cancer” arXiv preprint (2007).